A baraminologia nasceu morta, e já esta sepultada junto ao conceito de design Inteligente, vertente fanática do criacionismo que demonstrou ser um grande fiasco em 2005 por não sustentar-se no julgamento de Dover. O sistema de classificação dos criacionistas mostrou a incapacidade em favorecer qualquer evidência em favor de tipos básicos (baramins), pois nunca abraçou a totalidade das evidências, sempre pautado na convencionalidade e na seleção de características que favorecem uma verdade pré-concebida. Aqui, trataremos das falhas lógicas, estatísticas e conceituais por trás da baraminologia demonstrando o seu caráter mitológico: uma narrativa de caráter simbólico relacionada a uma dada cultura que busca explicar por meio de alegorias, folclores e personagens a origem das coisas; em especial na baraminologia, a criação dos seres vivos.
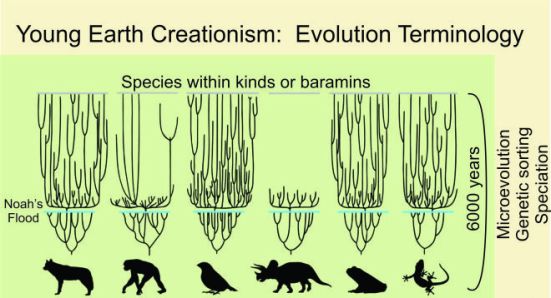
Esquema baraminológico onde cada grupo de animais é tratado de modo independente. O risco azul representa um suposto gargalo populacional promovido pelo dilúvio onde um casal de cada espécie representou um tipo básico (Baramin) que deu origem a diversidade atual de espécies. O esquema é pseudocientífico pois não tem respaldo empírico lógico e conceitual.
Baraminologia é um termo que emprega uma terminologia e metodologia não totalmente desconhecida pelos principais sistematas. O termo “baraminologia” vem de baramin, que foi construído a partir de palavras de raíz hebraica bara (criado) e min (tipo). Não é um conceito mencionado diretamente na bíblia, portanto, é uma construção fictícia para justificar um conjunto de falsas premissas criadas a partir de um relato teológico.
A ideia foi concebida pelo criacionista Frank L. Marsh em 1941. Baraminologia também tem sido referida como um “sistemática da descontinuidade”, pois não aceita a relação filogenética da totalidade da vida; e só aceita evolução se for rápida, a partir de ramos independentes da vida, sob um tempo geológico inferior a 6 mil anos.
Baramin é um grau taxonômico correspondente às “espécies criadas” forjado a partir de uma leitura do livro de Gênesis. Os proponentes do design inteligente que querem se livrar da aparência religiosa dada pelo criacionismo são promotores da baraminologia como uma forma de quanificar descontinuidades na árvore da vida e tendem a evitar o termo baramin, preferindo o termo “tipo básico” (Gishlick & College, 2006). Em resumo, baramin foi uma conjectura do hebraico inventada pelos criacionistas, e, um grupo deles prefere usar a tradução da palavra. A ideia é evitar implicações religiosas.
No Brasil esta tática não funciona, pois todas as comunidades de design inteligente no Brasil fazem propaganda do criacionismo, e os grupos de criacionismo fazem propaganda do design inteligente. De fato, todos os representantes de uma vertente, são exatamente os mesmos propagadores da outra. Desta forma, não há razão para separa-las, uma vez que os grupos e seus mesmos representantes bebem do mesmo corpo pseudocientífico (Veja mais em Design inteligente e Criacionismo – Darinha do mesmo saco (The Evolutionary Wedge).
O termo Baramin é usado como uma prova das ações de um designer ou criador especial segundo o criacionista ReMine. O objetivo é encontrar as “descontinuidades” na história da vida, ou dos limites de ancestralidade comum.
Os criacionistas usam muitos dos mesmos métodos que a maioria dos sistematas, a cladística da Análise método Padrão (anopa) e ferramentas para identificar as “lacunas”, ao invés das conexões na vida como a maioria dos sistematas fazem. De fato, o principal papel do criacionismo/design inteligente não é promover-se como uma ciência (porque já demonstrou claramente ser um discurso religioso fanático), mas criar um corpo (ainda que sustentado falsas premissas, distorção de artigos e conceitos falsos) de afirmações falaciosas para minar a credibilidade da teoria da evolução, mudanças climáticas e tudo que vai contra a crença de grupos fanáticos.
Quem nunca ouviu criacionistas afirmarem que a teoria da evolução foi plagiada, que a biologia não explica a origem de informação biológica, que os fósseis são forjados, que existe um complô mundial anti-cristão nas revistas científicas que impede a publicação de artigos de design inteligente, e até mesmo, que as mudanças climáticas são dados falsos e inventados por cientistas evolucionistas. Essas são só algumas das falácias mais propagadas, pois existem aquelas mais específicas.
Na análise estatística de caracteres para construir suas relações baraminológicas não é diferente. Existe um conjunto de dados selecionados que são jogados em uma matriz e que não seguem critérios matemáticos, ou que seguem modelos de relação que não fazem mais parte da sistemática atual.
Por exemplo; baraminologistas fazem uso de métodos fenéticos – que se baseiam em semelhanças gerais na aparência ou características gerais – e matrizes de distância de computação para grupo de táxons e produzem estatísticas com base no coeficiente correspondente de Sokal e Michener (1958). Eles vêem a fenética como útil para determinar lacunas biológicas. Coisa que os biólogos não adotam por não expressar um verdadeiro sistema de relação entre seres vivos, e também por ser um método ultrapassado, a mais de 60 anos.
Utilizar o máximo de características possíveis não reflete exatamente a ancestralidade comum (Futuyma, 1997). O problema é que os taxonomistas fenéticos agrupam por similaridade; então, segundo sua lógica, quanto mais semelhantes forem ás características, mais alto o nível que elas são agrupadas no fenograma. Desta forma, tal diagrama pode não representar a filogenia verdadeira por duas razões básicas: se um estado de caráter evoluiu independentemente em duas linhagens diferentes, ele indicará ancestralidade comum quando, de fato, as espécies não são derivadas de um ancestral comum imediato. Neste sentido, um morcego e um sabiá-galo-do-campo poderiam ser classificados como descendentes de um tipo básico comum na baraminologia. Pesquisadores interessados na reconstrução filogenética de uma espécie tendem a abandonar o método, pois considerando a evolução, existem sérios problemas em desconsiderar sinapomorfias, autopomorfias e homoplasias, por exemplo. Assim o método pela pura e simples semelhança estética de estruturas cria falsas correlações (Gishlick & College, 2006).
Neste ponto é interessante notar que corriqueiramente criacionistas dizem que semelhanças entre estruturas moleculares ou anatômico/morfológicas não são confiáveis para estabelecer relacionamentos evolutivos, mas eles utilizam as mesmas semelhanças estéticas de estruturas para estabelecer correlações baraminológicas. O problema é que a evolução biológica estabelece sim conexões confiáveis via homologias, apomorfias, inferências tiradas com base em dados empíricos, e não especulações retiradas de um livro religioso que inclusive, jamais disse qualquer coisa sobre tipos básicos (Gishlick & College, 2006).
Não é o simples fato de ser “esteticamente semelhante” que garante que duas espécies sejam relacionadas. A biologia evolutiva sabe disto, por isto tem métodos distintivos e não fenéticos de estabelecer relações filogenéticas. Não estamos mais na década de 50, a taxonomia evoluiu. A evolução consegue descobrir (na maioria das vezes) quando estruturas são homologas ou convergências evolutivas. Assim, os feneticistas são hipócritas por estabelecer conexões baraminológicas a partir de estruturas supostamente semelhantes, mas rejeitam as homologias (embora em alguns casos eles a usem sob outro nome e outra função) que de fato estabelecem conexões entre estruturas a partir da embriologia da evo-devo, e no nível genético e molecular, com genes Hox ou genes altamente conservados.
A outra razão pela qual a fenética da baraminologia é falsa tem a ver com as taxas diferenciais de evolução (Futuyma, 1997). O objetivo original do feneticista é estabelecer que o número de caracteres usados aumentasse e mais grupos fossem analisados, assim os resultados se repetiriam e eventualmente resultariam na classificação natural. Porém, muitas combinações podem ser feitas dependendo do programa utilizado para agrupamento dos táxons para a formação da matriz de dados. Esses programas oferecem uma série de dendogramas e ás vezes não há diferença significativa entre eles para que haja uma preferência entre os mesmos (Buchanan & Collard, 2007).
O aparecimento da escola fenética nos EUA, na década de 50 coincide com a construção das primeiras calculadores eletrônicas e computadores com grande capacidade de analisar dados. Os feneticistas se afastaram dos elementos evolutivos envolvidos na questão da diversidade para atender somente a elementos operacionais na taxonomia. A classificação fenética não esta interessada em exprimir a história filogenética de um grupo (Amorin, 2002), mas sim o interesse de grupos anti-ciência.
A ideia do uso da fenética (defendida por criacionistas como Robinson e Cavanaugh) era determinar relações intra-holobaramínica, bem como homoplasias (similaridades na forma não atribuídas a descendência comum) para grupos separados.
Na nomenclatura pseudocientífica da baraminologia, o holobaramin é um agrupamento que contém todos os organismos relacionados por descendência, não excluindo qualquer um. Um exemplo são os animais classificados como caninos. Eles são um holobaramin uma vez que lobos, coiotes, cães domesticados e outros canídeos são todos descendentes de dois tipos particulares levados a bordo da arca de Noé, e não há nenhuma outra criatura que seja geneticamente contínua com eles. Veremos mais adiante que este grupo biológico na verdade cria uma situação de desconforto na baraminologia ao não corresponder á premissa criacionista (Gishlick & College, 2006).
Para os criacionistas/design inteligente sinapomorfias (caracteres homólogos apomórficos compartilhados por dois ou mais táxons) são características que unem um holobaramin, e também é uma “descontinuidade” entre os grupos. Para eles (em especial Todd C. Wood da revista Baraminology), a sinapomorfia de um grupo sugere um evento criativo por Deus. Sim, curiosamente eles tentam catequizar o termo evolucionista sinapomorfia para uma versão cristã pró-Deus.
Outro problema que circunda a baraminologia é a definição precisa de “tipo”, uma vez que é um termo vago (como deixa claro o próprio criacionista Awbrey da Creation/Evolution). Eles fazem isto propositalmente, afinal, uma definição coerente irá permitir a descoberta dos tipos básicos criados, podendo finalmente, oferecer um cálculo do número de animais presentes sobre a arca, para os criacionistas da Terra jovem. Este cálculo assusta os criacionistas, já que poderia criar números altíssimos, superiores ao que a suposta arca poderia abarcar (Gishlick & College, 2006).
Há outros termos dentro da nomenclatura pseudocientífica da baraminologia; monobaramin, apobaramin e polibaramin.
Monobaramin é um grupo contendo somente organismos relacionados por descendência comum, mas não necessariamente todos eles. É um grupo ad hoc de organismos que compartilham descendência comum. Por exemplo, lobos são um monobaramin do holobaramin canino. Apobaramin é um grupo consistindo da totalidade de pelo menos um holobaramin. Um grupo consistindo de ambos humanos e caninos é apobaramínico já que ambos os membros são holobaramins. Um polibaramin é definido como um grupo consistindo de parte de pelo menos dois holobaramins. É um grupo ad hoc de organismos onde pelo menos dois dos membros devem ser não relacionados. Por exemplo: Humanos, lobos e um pato são um grupo polibaramínico (Gishlick & College, 2006).
O objetivo final dos criacionistas/design inteligente é pegar polibaramins e dividi-los em seus componentes monobaramínicos e seus respectivos holobaramins. Cada holobaramin pode então ser colocado em apobaramins de animais estruturalmente semelhantes.
Baraminologistas sugerem que isto é útil para falar sobre apobaramins porque os holobaramins têm muitas semelhanças que cruzam fronteiras holobaramínicas. Apobaramins são considerados úteis para o estudo de grupos maiores de animais morfologicamente semelhantes. Ora, os seres humanos deveriam ser comparados com o grupo mais estruturalmente e funcionalmente semelhantes a eles: os grandes primatas. Assim, se esses dois grupos não são filogeneticamente relacionados como pressupõem os baraminologistas, porque não apresentam os dados comparados?
Uma vez que os holobaramins tenham sido determinados, as relações filogenéticas dos membros em grupo de um baramin poderiam ser trabalhadas. No entanto, baraminologistas não acham que a simples exibição dessas relações em forma de árvores seria útil. Eles produzem as árvores, mas fazem algo diferente do que simplesmente produzi-las para descrever relacionamentos; eles sugerem modos alternativos de relação baseado em redes, treliças, ou parcelas de projeção de reconhecimento de padrões para estabelecer as relações (Gishlick & College, 2006).
Membros de um holobaramin poderiam ser resultado de uma triagem para a prole de diferentes genes (ou de DNA no geral) de organismos parentais. Para eles, desde o tempo da criação poderia ter havido algumas modificações hereditárias do DNA (mutações), que foram passadas para descendências divergentes.
Curiosamente os baraminologistas (assim como os evolucionistas) afirmam evitar pensar na concepção “essencialista”. O que é estranho, dado que as suas limitações são sugestionadas a “tipos básicos”, ou seja, tipos essenciais na qual todos os outros se desenvolveram. Eles afirmam isto porque reconhecem que as características baraminológicas podem ser perdidas através da variação dentro de uma espécie. E embora o termo/classificação baraminológica não esteja na bíblia, a definição essencialista que ele tentam evitar esta presente em Gênesis 1:11-12. E toma partido de que o tipo básico é a espécie. Se a intenção é interpretar a bíblia literalmente, eis a definição “correta” do termo:
11-E disse Deus: Produza a terra erva verde, erva que dê semente, árvore frutífera que dê fruto segundo a sua espécie, cuja semente está nela sobre a terra; e assim foi.
12-E a terra produziu erva, erva dando semente conforme a sua espécie, e a árvore frutífera, cuja semente está nela conforme a sua espécie; e viu Deus que era bom.
Em resumo, o criacionismo criou termos bíblicos que não existem, e discursa contra o essencialismo da bíblia, ainda que seja favorável a ele, nem que para isto seja necessários criar um modelo de classificação próprio, pautado em falácias feneticistas.
A busca pelas Descontinuidades
A ideia dos baraminologistas é seguir os dados para descobrir descontinuidades nos relacionamentos filogenéticos apresentados pela teoria da evolução. Em suas análises, frequentemente e explicitamente critérios bíblicos são usados quando descontinuidades não são encontradas entre os grupos que “deveriam ser” descontínuos. Este é o caso dos seres humanos e chimpanzés em que encontrados nos textos de Robinson e Cavanaugh postados pela Creation Research Society Quarterly; organização cristã promotora do criacionismo e design inteligente. O criacionista Tood C. Wood afirma explicitamente em “An evaluation of lineages and trajectories as baraminological membership criteria” que os critérios bíblicos são primordiais e que descontinuidades são pressupostas por causa dos eventos de criação separadas mencionadas na Bíblia (Gishlick & College, 2006). Assim, baramins podem ser definidos em uma série de critérios, de tal modo que o que importa é a validação da conclusão pré-concebida; a criação é obrigatoriamente divina. Fica claro então que a apresentação baraminológica não é científica, mas religiosa, seja pelo próprio nome da taxonomia criacionista ou pelas declarações fanáticas de seus representantes.
Usar uma série de critérios de adesão para determinar os limites holobaramínicos e todas as medidas é considerado um método falível, e, mesmo assim, os defensores argumentam que vários critérios devem ser empregados na tentativa de diagnosticar um holobaramin (Wood Cavanaugh e muitos outros partilham desta ideia). Assim, o critério criacionista é utilizar tudo quanto é critério, discurso e máximas para validar a ideia pré-concebida de criação e fomentar um preconceito e discurso anti-científico contra a evolução.
Para medir a distância baraminológica entre duas espécies eles empregam uma ampla gama de métodos para descobrir ou forjar lacunas morfológicas, recorrendo a métodos tradicionais como a cladística e a fenética para desenvolver seus próprios métodos.
Na apuração de tais unidades, são usadas medidas de consistência cladística tradicionais e medidas fenéticas de distância, juntamente com outros métodos estatísticos multivariados. Incluindo critérios morfológicos na identificação de morfologias, órgãos, vias metabólicas, processos celulares, ou funções que são exclusivas de um grupo (por vezes consideradas sinapomorfias) e, portanto, supostamente sugestivas a origens distintas. É uma distorção dos dados originais para suportar uma premissa teológica (Gishlick & College, 2006).
O baraminologistas começam com uma classificação filogenética hipotética (o que é irônico, dado sua negação das relações filogenéticas em grande escala), assumindo que o grupo conteria grupos verdadeiramente holobaramínicos. Animais de grupos taxonômicos intimamente aliados são agrupados e os grupos de vizinhos mais próximos são selecionados como grupos externos para comparabilidade biológica. É como criar um grupo de borboletas e usar como grupo externo de comparação outro grupo de borboletas e no final estabelecer que o grupo de borboletas foi criado divinamente em apenas um grupo, isolado descontínuo das mariposas, ou estes, de trichopteros (ordem de insetos evolutivamente mais próxima das borboletas/mariposas).
Baraminologistas também usam métodos cladísticos. Eles calculam árvores usando software de cladística tradicional. Grupos com altas correlações dentro de um conjunto de dados bootstrap são considerados potencialmente holobaramínicos e, em seguida, são testados para validar a descontinuidade filogenética dos seus subgrupos. Altos níveis de homoplasia também são considrados indicativos de baramins separados, que baraminologistas propõem serem resultado de criações separadas usando como referência a bíblia.
Baraminologistas consideraram também que homologias existem dentro holobaramins; e homoplasias são características compartilhadas entre holobaramins (como defende o criacionista Wise da Creation ex Nihilo Technical Journal). Eles também assumem que certo nível de homoplasia delineia uma descontinuidade filogenética, mas nenhum critério é utilizado para definir porque eles tratam tais características como evidência de holobaramins. Geralmente partem do pressuposto, não testado de que os organismos são muito bem projetados. (Veja As falhas de projeto mais infelizes do corpo humano)
A fim de determinar o grau de homoplasia, os baraminologistas calculam um “índice de Homoplasia” (IH) o equivalente de 1-IC de análise filogenética tradicional (Kluge e Farris 1969). Se o IH é alto, então são os baramins preferidos são separados. Se o IH é elevada dentro de um holobaramin, propõe-se que seja o resultado de um “espalhamento do gene” a partir de um antepassado complexo através de hibridação.
O problema é que estatisticamente não há nenhuma medida específica de que grau de IH representa baramins separadas e nenhuma explicação de por que isso se encontra alto dentro de um holobaramin diagnosticado, o que não prova que ele é realmente um apobaramin. Isso significa que nem os baraminologistas tem um critério coerente de seleção.
Há um índice 0 homoplasia entre humanos e macacos em seu conjunto de dados, o que sugere ser uma medida imperfeita por cauda dos critérios em algumas circunstâncias; aqueles em que a análise não produz a resposta que eles querem. Por isso, eles alertam sobre como usar o IH como critério; é preciso distorcer os dados quando compara-se humanos e primatas, pois na condição forjada deles o resultado colocaria humanos e chimpanzés no mesmo grupo, e neste caso, a analise precisa de um ad hoc para separar as duas espécies holobaramínicas relacionadas. Isto significa que mesmo dentro do sistema de classificação informal deles, o homem e os outros grandes primatas são agrupados. É preciso alterar a matriz de dados e criar manobras alternativas ad hoc e fugir da base estatística e cladística que relaciona homem e outros primatas de grande porte. É uma alteração do critério que eles fazem descaradamente para evitar o resultado estatístico de relacionamento evolutivo entre as espécies (Gishlick & College, 2006).
Quando usam dados moleculares com a desculpa de que oferecem a oportunidade de procurar descontinuidades no nível “fundamental” da vida a historia não é diferente.
O raciocínio é idêntico ao de sistematas tradicionais, mas a baraminologia utiliza alguns dos mesmos dados e técnicas analíticas, entretanto, acreditam que todos os holobaramins passaram por um gargalo grave, que eles consideram ser a data do dilúvio de Noé. Assim, o DNA mitocondrial deve ser uma ferramenta sistemática ideal (segundo o criacionista Robinson da Creation Research Society Quarterly). Isso por si só já deixa claro que apesar de não gostar de usar o termo baraminologia para não evidenciar um caráter religioso, as premissas são claramente religiosas.
Os baraminologistas baixam sequências de bases de dados on-line, tais como GenBank (que é a base de dados da evolução onde estão muitas sequencias evolutivamente relacionadas) e usam métodos típicos de alinhamento filogenéticos. Eles compararam sequências de diferença percentuais em comparação com sequência de classificação taxonômica, usam estimativas de distância por parcimônia para construir agrupamentos e, finalmente, avaliam esses agrupamentos por métodos de bootstrap, calibrando com a bíblia. O único problema é que eles usam uma taxa de diversificação que eles acreditam que são “filogeneticamente distintas” para grupos externos em suas análises moleculares (Gishlick & College, 2006).
Outra forma de avaliação baraminológica é com base nos critérios ecológicos que foram propostas pela primeira vez por Wise, que argumentou que as diferenças ecológicas e tróficas refletiam a origem ou grupos separados. Ele baseou-se na ideia de que a classificação taxonômica de família refletiu espécies criadas e suas observações é que as famílias tendem a conter os animais com ecologias semelhantes e níveis tróficos. Assim, diferentes características ecológicas ou tróficas deve delinear baramins separados. Sugerem que este critério seria mais útil para organismos unicelulares, citando os radicalmente diferentes ecologias e metabolismos celulares encontrados em bactérias e archaeas, que, segundo eles, sugerem origens distintas. Citam semelhanças entre arqueas, eucariotos e procariotos e que o registro estratigráfico não é o reflexo da história ancestral dos seres vivos, mas da deposição durante o dilúvio de Noé. Em outras palavras, para eles a formação dos imensos cânions (Veja 1 2 e 3), montanhas e formações geológicas são frutos de 40 dias e noite de chuva e um ano de decantação; tempo que demorou para a água do dilúvio escorrer para um lugar na qual nenhum criacionista soube explicar qual é, nem de onde veio tal volume de água (Veja Geologia do Dilúvio – Afundando a arca de Noé). Isto remonta os tempos de discussão do Immanuel Velikovsky (Gould, 1999).
De fato, com essas preposições baraminológicas, a posição estratigráfica de organismos deve ser irrelevante sob este modelo. Para o criacionista Marsh, a ideia é que podem ser estabelecidos os limites de um baramin para um grupo de organismos pela sua capacidade em hibridizar. Não importa se a hibridação é natural, ou se a descendência é fértil, mas apenas que a hibridação é possível através de alguns meios, incluindo a inseminação artificial. Baraminologistas coletam dados e catálogo híbridos e complementam com a hibridação de sua própria autoria e aplicam critérios que lhes interessa.
Existem dois tipos de critérios bíblicos; o primeiro se a Bíblia faz referência especificamente ao baramin como especialmente criado, e o segundo, se a Bíblia indica que foi especialmente criado (de acordo com o criacionista Wood).
Primeiramente os baraminologistas procuram qualquer identificação de uma baramin em textos bíblicos. O criacionista Robinson sugeriu tartarugas, que são identificados em Levíticos 11-29:31.
29-Estes também vos serão por imundos entre os répteis que se arrastam sobre a terra; a doninha, e o rato, e a tartaruga segundo a sua espécie,
30-E o ouriço cacheiro, e o lagarto, e a lagartixa, e a lesma e a toupeira.
31-Estes vos serão por imundos dentre todos os répteis; qualquer que os tocar, estando eles mortos, será imundo até à tarde.
Em segundo lugar, eles determinam se o animal é “limpo” ou “impuro”, determinando quantos pares foram trazidos na arca. As tartarugas são determinadas como animais impuros, ou imundos; assim, apenas um par de cada baramin de tartaruga teria sido necessário. Robinson também sugere que as tartarugas marinhas não teriam estado na arca. No caso dos seres humanos e primatas, Robinson e seu colega Cavanaugh concluem que, apesar de outros critérios não poderem separar os seres humanos e primatas, os critérios bíblicos afirmam especificamente que os seres humanos formam um baramin separado, já que foi criado a imagem e semelhança de Deus.
Para ajudar na busca pela descontinuidade, baraminologistas desenvolveram duas metodologias; anopa e BDIST.
Em anopa eles consideram que a “verdadeira” linhagem é parte do critério morfológico e estratigráfico. A ideia por trás deste pressuposto é que os organismos poderiam ser representados como pontos discretos num morfoespaço tridimensional. Se organismos podem ser ligados por uma linhagem contínua no morfoespaço, então eles podem ser considerados como parte do mesma baramin. A anopa é uma ferramenta estatística desenvolvida pelo criacionista Cavanaugh para determinar se tais linhagens existem. O próprio Cavanaugh afirma que este método é útil para investigações tridimensionais de dados morfológicos quantitativamente; no entanto, parece que pouco diferem da análise de componentes principais, com uma exibição gráfica fantasiosa com medidas estatisticamente suspeitas. Baraminologistas também desenvolveram o seu próprio software de análise; o BDIST, que calcula o “coeficiente de distância baramínica”. Esta distância representa a porcentagem de caracteres de dois taxa em comum. Se não houver uma “cadeia positiva de correlações de distância baramínica significativa” que conectam todos os taxa, então eles são monobaramínicos. Basicamente, BDIST calcula uma matriz fenética de distância (Gishlick & College, 2006).
A metodologia BDIST não tem sido amplamente aplicada, e não há evidências de que os efeitos algoritmos de grandes conjuntos de dados sejam reais. Os baraminologistas parecem aplicar-se no método fenético de idade, sem realmente estudar como ele funciona. Em nenhum dos casos ele realmente distingue entre dois baramins.
Vejamos um exemplo; os baraminologistas determinaram um status holobaramínico para um grupo, como é o caso de felídeos e fósseis e recentes eqüídeos. Neles, não há retorno de resultados favoráveis para baraminologistas, e então, outros critérios são aplicados para atingir o resultado desejado, partindo da conclusão pré-concebida criando um método que respalde A “Verdade” absoluta. Este também foi o caso para os seres humanos e primatas, onde BDIST não apresenta uma separação para essas espécies. Os autores empregam métodos ad hoc como os “critérios ecológicos” e o critério bíblico para atingir baramins separados, apesar de não discutir os “critérios bíblicos” (Gishlick & College, 2006).
Somente a análise baraminológica de eqüídeos (feita por Cavanaugh em 2003) incluía tanto fósseis e taxa vivo, e nenhuma das outras análises continha conjuntos de dados. Portanto, este estudo serviu de modelo para ver como eles iriam investigar fósseis e conjuntos de dados morfológicos recentes. A estratégia criacionista vem da reinterpretação de dados de fósseis com datadas em milhões de anos que passam a ser ignorados em sua totalidade e servem para serem distorcidos em uma cosmovisão teológica.
Evander (1989) coletou e analisou dados de cavalos fósseis. Cavanaugh, o baraminologista, removeu os dados em falta por recodificação para que os dados “desconhecidos” fossem codificados como 0, ausência como 1, derivação parcial como 2, e presença como 3 e 4. Os dados desconhecidos para um valor específico que podem ser utilizados na análise faz com que o conjunto de dados de todos os caracteres apresente um valor falso e um aumento da quantidade de variação morfológica através da atribuição de um valor numérico a algo desconhecido (Gishlick & College, 2006).
Toda esta manobra pseudomatemática e estatística da matriz de dados foi forjada para acelerar a taxa de especiação e justificar como em tão pouco tempo poderia um tipo básico dar origem a diversidade de equídeos atual. Tal resultado foi um fiasco.
O software BDIST, conforme configurado, apresenta qualquer caractere com menos do que “95% de relevância”. Por que este limiar foi escolhido não é explicado, nem como ele é determinado. A relevância parece ser determinada pela percentagem de dados em falta para uma característica em relação ao número total de táxons. Isso faz com que a aplicação deste método para conjuntos de dados fósseis inúteis e falsos. O uso de dados “requentados” (recodificação dados) como valorizados, como pressupôs Cavanaugh em 2003 deram um efeito mensurável distorcido sobre os resultados, especialmente se a quantidade de dados perdidos apresentarem uma proporção maior do conjunto de dados presentes.
Quando as distâncias baramínicas relativas são comparadas com as relações filogenéticas produzidas pelos conjuntos de dados, o resultado global é uma tendência constante e gradual na diminuição da distância baramínica em relação à filogenia. Assim, sem mudanças significativas dentro distância baramínica dos conjuntos de dados, foi possível concluir que os dinossauros e aves pertencem ao mesmo holobaramin. Isso faz sentido em um contexto evolutivo: as características mais transitórias encontram-se em um conjunto de organismos relacionados, pois será menor a distância relativa entre dois taxa.
O biólogo evolucionário Phil Senter fez um teste interessante e publicou na revista científica Journal of Evolutionary Biology. Ele empregou o método CMDS (Classic MultiDimensional Scaling), usado pelos baraminologistas para visualizar continuidade ou descontinuidades morfológica entre grupos taxonômicos (Senter, 2010) em dinossauros terópodes coelurosaurianos. Como os criacionistas consideram a continuidade como sendo evidência de parentesco genético e a descontinuidade como evidência de criação independente, ao aplicar o método CMDS em dinossauros e as aves, o programa mostrou uma continuidade entre os Archaeopterix (aves primitiva descendente dos dinossauros) e outros dinossauros terópodes não-avianos. Em resumo, o Archaeopterix foi visto como uma forma de transição evolutiva, ou continuidade, pelo próprio método criacionista. Mesmo os métodos criacionistas, quando aplicados de maneira rigorosa e consistente, não apoiam a maioria de suas crenças e apontam para a ancestralidade comum. Razão pela qual distorcem e omitem dados das matrizes, para criar analises estatísticas frágeis e superficiais, criando descontinuidades e justificando-as como criações divinas. Esta é a essência e a intenção dos baraminologistas; usar pseudomatemática para justificar a existência de um designer.
Ainda sim os criacionistas alegaram que o método usado pelo biólogo Senter não era o ideal, pois apenas fornecia um método de visualização de dados e não uma “inferência cientifica”, sendo apenas um método de “correlação de táxons”. Wood, citado anteriormente, fez então uma contra-análise deste tipo e chegou a um resultado diferente ao de Senter no caso dos dinossauros, encontrando 3 baramins ao invés de apenas um (Senter, 2011). Como uma grande quantidade de fósseis de transição foram sendo encontradas nos últimos anos, naturalmente começaram a surgir ás ligações entre os diferentes taxa de dinossauros (torando a baraminologia cada vez mais exposta a farsas estatísticas), aumentando a continuidade morfológica dentro da ordem Dinosauria e corroborando a afirmação feita pelos biólogos evolutivos e paleontólogos que todos os dinossauros filogeneticamente relacionados descendendo de um ancestral comum (Senter, 2011).
Senter então resolveu usar o método baraminológico de “correlação de táxons”, usado por Wood para avaliar essa questão diante dos novos achados que foram incorporadas e uma série de correções e continuidades que foram feitas as matrizes de dados usadas por Senter resultando em um segundo artigo mostrando as continuidades onde Wood não havia encontrado. Não foi necessário incorporar descobertas de novos fósseis, pois a correção dos dados e a inclusão de outros espécimes já conhecidos (que foram negligenciados), mas que não estavam na matriz original fecharam descontinuidades.
Criacionistas podem ainda sim alegar que existem certas descontinuidades, e de fato existe, por isto a evolução busca mais fósseis e dados. Entretanto, ainda sim não é vantajoso para eles recorrerem a este tipo de argumento da ignorância e do ‘Deus das lacunas’, por essa razão, os baraminologistas andam meio cautelosos e sumidos com seus textos. Grande parte do público que recorre a baraminologia para tentar corroborar suas crenças é formado por leigos, que aceitam o método baraminológico sem compreende-los, e sem saber das falhas graves de seus métodos. Por fim, acabam replicando cegamente um método que não se sustenta logicamente, e carece de dados empíricos. São replicadores de falácias e pseudomatemátcas.
O simples fato de existir lacunas não pode ser usado como argumento para mostra que a evolução nunca aconteceu, até porque, um arcabouço enorme de evidências empíricas estabelecem a realidade do fenômeno para a comunidade científica que cotidianamente acumula dados e evidências aprimorando nosso conhecimento de como este fenômeno se deu e ainda se dá, expressando os detalhes de seus mecanismos.
Em conclusão, o programa de software empregado pelos baraminologistas sugere que o método não funciona, pois é forjado em bases estatísticas falsas, sintetizadas a fim de contemplar uma afirmativa pré-concebida, distorcendo o método para se encaixar em uma verdade dita absoluta, seja para criacionistas seja para proponentes do design inteligente, que historicamente, na teoria e na prática, são exatamente o mesmo movimento anti-ciência. A supremacia dos critérios bíblicos é explicitamente admitida, razão pela qual os textos são publicados em revistas religiosas já que ao há critério estatístico que passe pelo crivo de uma revista científica.
Outras críticas
Outra critica que é comumente feita quando se discute a impossibilidade da baraminologia vem da necessidade de encontrar o fóssil do “tipo básico”. Geralmente os fósseis são encaixados dentro de um relacionamento baraminológico que descende de um misterioso tipo básico que nunca é encontrado em registro fóssil. De fato, todos os indivíduos encontrados pertencem a um grupo já conhecido. Tomemos um animal como exemplo; os leões que são encontrados em registros egípcios e gregos a mais de 5 e 3 mil anos. Neste período já existiam os leões modernos e outras espécies hoje extintas, mas que sabidamente eram leões.
Sansão, o herói da Bíblia conhecido pela sua força, supostamente matou um leão com suas mãos (Juízes 14:5-9). Ele nasceu em 1.200 a.c. Ou seja, já existiam leões modernos naquela época. Daniel é outro personagem bíblico conhecido por ter entrado em uma cova onde havia leões no século V a.c. Tal animal poderia representar o extinto leão-das-cavernas que ainda sim era um leão e não um tipo básico. Em todas as passagens bíblicas e não bíblicas notamos o relato de leões, mas não de seus baramins, ou sobre processos de especiação rápidos o suficiente para contemplar a visão baraminológica.
Ao mesmo tempo, notamos que as espécies antigas de leões foram extintas não pelo episódio do dilúvio, mas pela própria dominação humana. Fragmentos ósseos encontrados na Espanha, Itália e Grécia são atribuídos tanto ao leão moderno quanto ao leão-das-cavernas. No Holoceno Superior, há restos ósseos de leões datados do Neolítico, entre 3.500 e 500 a.c. Na Grécia, os leões extinguiram-se no Peloponeso durante o final da era micênica em 1.400 a.c. (Schnitzler, 2011). Heródoto menciona que leões viviam nas planícies de Nestos, tendo atacado as caravanas de camelos do rei persa Xerxes I em sua marcha através da Trácia em 480 a.c. Aristóteles relatou que eles eram raros por volta de 300 a,c (Schaller, 1972). Sendo completamente extintos nas montanhas do norte da Grécia no primeiro século d.c (Schnitzler, 2011).
Notamos então que nenhum episódio teria dado origem a diversas espécies de leões, afinal, elas já existiam muito tempo antes. Nenhum tipo básico semelhante a leões é relatado, não há evidências genéticas de explosão de diversidade de leões nos últimos 4 mil anos. Há somente relatos de extinção por atividade antrópica, e não por dilúvio.
Observemos a proposta da evolução biológica.
De fato, a origem do gênero Panthera é datada de 6,4 milhões de anos. O registro fóssil mais antigo de um leão é marca 2,8 milhões de anos. A análise genética de leões que ainda vivem e de espécimes de museu confirmam que o ancestral comum mais recente dos leões modernos viveu por volta de 124 mil anos atrás. Os leões modernos evoluíram em dois grupos; na África Oriental e Austral, a outra inclui leões na África Central e Ocidental, e na Índia segundo artigo publicado na revista científica BMC Evolutionary Biology. De acordo com este estudo nesse tempo, as florestas tropicais expandiram-se através da zona equatorial da África e região do Sara virou-se para savana. Os leões que vivem no sul e leste do continente se separaram por afastamento territorial dos que vivem no oeste e norte. As diferenças genéticas entre estes dois grupos de leões permanecem até hoje. Cerca de 51 mil anos atrás, o continente seco e o Sahara expandiu, cortando as populações de leões no oeste daqueles no norte. Ao mesmo tempo, os leões no oeste expandiram para a África Central, que se tornou mais habitável. Desde então, grandes rios da África, incluindo o Nilo e Níger, têm ajudado a manter esses leões afastados.
Outro detalhe revelado apenas pelo estudo do DNA antigo de espécimes, é que os leões modernos começaram o êxodo para fora da África a apenas 21 mil anos atrás. No final do Pleistoceno, leões deixaram o Norte de África, eventualmente, chegando tão longe, como a Índia. Mais tarde, apenas 5.000 anos atrás, outro grupo de leões deixou o continente, atingindo o que é hoje o Irã, no Oriente Médio. Estes leões estão agora extintos.
Para piorar a situação dos baraminologistas o leão-das-cavernas era representado em pinturas rupestres atribuídas aos primeiros seres humanos da Europa, o que demonstra que sobreviveu até ao Holocêno. Os desenhos de felinos, claramente de leões, são datados de 32 mil anos da caverna Chauvet (França) e demonstra juba e um tufo de pêlos na ponta da cauda.

Mauricio Antón & Alan Turner: Os Big Cats e registros em Chauvet. Columbia University Press, 2000,
O leão-americano ou megaleão (Panthera leo atrox ou Panthera atrox) é um felino extinto que viveu na América do Norte durante o Pleistocêno e extinguiu há cerca de 10 mil anos. A espécie ainda tem classificação discutível, mas geralmente é considerada como um sub-tipo do leão moderno (Panthera leo) e próxima do leão-das-cavernas que viveu na Europa durante o mesmo período. É também chamado de leão norte-americano, Naegele, onça gigante ou leão das cavernas americano. Ele foi endêmico da América do Norte durante e sua origem data pelo menos 340 mil anos. Um dos primeiros leões conhecidos nas Américas sul foi encontrado ainda no Alasca, do Sangamonian Stage, o último interglacial. Depois disso, o leão americano espalhou-se amplamente de Alberta para Maryland, e até o sul de Chiapas, no México (Marisol & Carbot-Chanona, 2009).
Esses achados demonstram que a história dos felinos em geral não pode ser traçada a partir de um tipo básico que evolui rapidamente e posteriormente ao conto de Noé recolonizou continentes uma vez que já estavam separados em apenas 6 mil anos.
Não existiu um tipo básico para leões, que o distingue de tigres, onças ou leopardos. De fato a linhagem Panthera é extremamente relacionada ao gênero Neofelis. Ambos os grupos são relacionados com o grupo dos gatos do mato. Todos os felinos descendem de um ancestral comum, o Pseudaelurus que viveu na Ásia até pouco mais de 11 milhões de anos atrás e se divergiu nos grupos de felinos que conhecemos hoje. O primeiro grupo de felinos surgiu a 10 milhões de anos e deu origem aos indivíduos do grupo pantera. E antes do Pseudaeurus outros felideos já existiam. Alguns felinos como o dente-de-sabre já existissem a mais de 35 milhões de anos.
Análise filogenética revela a ocorrência de apenas três clados distintos de leões: o leão moderno, o leão-das-cavernas da Eurásia e Beríngia, e o leão-das-cavernas-americano, que forma uma população separada ao sul das geleiras pleistocênicas. Um táxon denominado vereschchagini se mostrou indistinguível do spelaea através da análise genética de DNA antigo, sendo considerado como sinônimo do leão-das-cavernas-europeu (Barnett, et al, 2009).
A história dos leões assim como de qualquer outro grupo não pode ser segregada em linhagens distintas, mesmo porque o termo “tipo” é muito vago. As descontinuidades não existem, seja dentro do grupo dos felídeos, seja em níveis maiores.
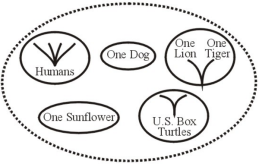
Neste esquema baraminológico de Frair considera tigres e leões como fruto do mesmo baramin. Os outros felinos tem outros baramins ou são fruto deste baramin? Como tantos felinos poderiam evoluir de um mesmo baramin em alguns milhares de anos? Nenhum criacionista propôs mecanismos que expliquem essas questões.
Se há aceitação de que espécies descendem, ou emergem de baramins, isto é tecnicamente a aceitação da macroevolução, afinal, leões e tigres não são as mesmas espécies, e se apresentam um ancestral em comum, são exemplares claros de evolução. Neste sentido, é muito mais coerente á aceitação da teoria da evolução como um todo, do que aceitar segundo a conveniência de uma crença pautada em cima de ad hocs com metodologias estatísticas falhas. Isto porque se a definição de baramins for ampla (em gêneros, famílias ou ordens, por exemplo) torna a aceitação de um processo de descendência com modificação e divergência de linhagens.
A falta de definição de um baramin é um peso muito grande contra a baraminologia. E mesmo quando é dada cria situações conflitantes. Vejamos um exemplo.
Imaginemos um baramin para um cão, que certamente abarcará o cão doméstico (Canis lupus familiaris), bem como o lobo (Canis lupus lycaon), o dingo (Canis lupus dingo) e o coiote (Canis latrans). O problema é que outros membros do gênero Canis, como por exemplo os chacais, são normalmente incluídos também. Mas nem todas as espécies de chacais podem hibridizar com cães. Isto fere o conceito de baramin. Se pegarmos ursos, guaxinins e doninhas; veremos que são classificados na subordem Canoidea. Criacionistas não aceitam que são o mesmo baramin ainda que cumpram todas os requisitos básicos. Dentro do mesmo baramin criacionistas nem que ursos e cães possam se originar em apenas 6 mil anos de especiação baraminológica. Nem eles aceitam as suas propostas. Eles tentam definir um baramin para cães e um para ursos, ainda que biologicamente apresentem evidências que os relacionem. Veja você mesmo aqui.
Se critérios são criados a partir da semelhança morfológica, porque rejeitam a classificação de Canoidea? A resposta é que a aceitação é condicional ao que eles querem ver como baramin, e se guiam por critérios bíblicos e não factuais. Por exemplo; qual seria o baramin de répteis? Se há um baramin para répteis, ele tem de receber desde as serpentes até dinossauros, ictiossauros, pterosauros e anfisbenas. Deveria abarcar toda a biodiversidade do grupo como resultado de 6 mil anos de evolução.
Outro ponto é; chamar de baramin não muda o fato de que um grupo dá origem a outro, e isto é em essência um modelo evolutivo darwiniano. Em síntese, quando eles negam a evolução e a ancestralidade comum, eles aceitam taxas de modificação evolutivas que não tem respaldo empírico nem estatístico ou mesmo lógico. Criam um conjunto de premissas incompatíveis com o que a biologia sabe sobre ecologia, genética em cada baramin.
Uma das críticas a baraminologia refere-se ao pós-Éden. Se os animais eram “herbívoros” em sua maioria antes do dilúvio, com a condição de expulsão do jardim divino não haveria pressão e portanto não haveria especiação e os tipos básicos seriam os mesmos até hoje, uma vez que passaram somente 6 mil anos da criação, e 4 mil após dilúvio.
Outro questionamento simples é; se há um tipo básico para primatas porque este tipo básico ancestral não diferenciou os primatas de grande porte de nós? Seria o homem e o chimpanzé convergências baramínicas apresentando 99% de semelhança de DNA, número de cromossomos próximos, com redes de genes semelhantes por puro fruto da criação intencional? Seria a semelhança entre os dois grupos fruto do acaso ou de um designer brincalhão?
A baraminologia pós-mito diluviano.
Infelizmente para os criacionistas a questão do dilúvio fica restrita ao campo da teologia e dos mitos. Razão pela qual, esta será a última vez que o NetNature aborda tal questão, já que não faz parte do conteúdo científico e porque grande parte das críticas feitas ao dilúvio estão centradas no texto Geologia do Dilúvio – Afundando a arca de Noé.
Tomemos como referência a primeira figura do presente texto, um relacionamento baraminológico. Supostamente, algumas linhagens escaparam da punição diluviana proporcionada pelo Criador, graças ao bom Noé (ou ao “malvado” Noé, representado por Russell Crowe na versão judaica do mito). E como vimos acima, há relatos confiáveis de muitos animais datados em milhares de anos; como o exemplo dado dos leões.
Se a diversidade da vida teve um re-início com o dilúvio seria esperado encontrar fósseis dos tipos básicos anteriores ao dilúvio, o que não ocorre. Seria de se esperar também uma taxa de especiação extremamente alta após o dilúvio, diante de um tempo de especiação bem menor em relação ao momento da criação de Gênesis 1:11-12. Mesmo com a redução do contingente animal dentro da arca, problemas alimentares, predação seria evidentes e principalmente, o pós-dilúvio no que diz respeito a dispersão dos animais em torno do globo e a diferenciação do tipo básico rápida e que hoje estaria cessada. Destacamos também que não há relato nos povos antigos sobre a explosão de diversidade de animais que eram de um único tipo e passaram a ser vários.
Como explicar o fato de tamanduás e capivaras existirem somente na América do sul. Ou porque há 3 espécies de antas na América do Sul/Central e uma única isolada no Vietnam. Como explicar o endemismo do castor no país do criacionismo?
Isto ocorre porque o relato do dilúvio, assim como a baraminologia, se respaldam sobre um relato teológico, fundado em mitos antigos. O relato bíblico não é factual, mas sim um discurso de devoção, que não precisa se sujeitar a verdade e sim aos mitos ou interesses de grupos específicos. Ele foi inspirado em contos mais antigos que a tradição judaico-cristã e portanto, deve ser encarado como tal, como veremos.
O conto sumério de Gilgamesh (2300-2000 a.c) é registrado em fragmentos de textos traduzidos pelo povo Hitita e Hurrita e é também retratado em grego por Babiloniaka de Berose, um sacerdote que difundia a cultura helenística no século III a.c (História viva, 2015). No conto babilônico, o deus supremo Enlil queria destruir a humanidade, mas Ea, uma divindade clemente e precavida advertiu tal intenção de Enlil ao seu fiel escudeiro humano Uta-Napishtim-Ruqu. Ea ordenou que ele fizesse um barco de 120 côvados para salvar sua família, amigos, riquezas, seus mestres e obras antes do dilúvio. Ele o fez, lacrou a arca com betume e salvou-se. Tal dilúvio apavorou até outros deuses do panteão. Após o dilúvio Uta-Napishtim-Ruqu soltou uma pomba, e posteriormente uma andorinha, na qual regressaram a embarcação. Após alguns dias, liberou um corvo que nunca regressou. Então, sua arca repousou sobre o monte Niçir e ao sair preparou um banquete em oferenda aos deuses. Enlil notou a sobrevivência do homem e furioso cobrou explicações de Ea, que se justificou afirmando que sem os homens não haveria refeições e oferendas aos deuses destacando os motivos pelos quais salvou a humanidade (História viva, 2015).
No conto acadiano (1750-1600 a.c), o poema de Atrahasis (que apresenta poucos fragmentos) relata a mesma história, com o deus Enlil, porém, a motivação de tal dilúvio veio porque ó homem era muito barulhento e não deixava os deuses descansar em paz. No conto sumeriano, o rei Ziuzudra é avisado em um sonho por Enki (que representa o deus Ea) que um dilúvio de 7 dias e 7 noites viria, e ele ficou encarregado de criar uma arca para salvar a humanidade. Após o dilúvio, Ziuzidra ofertou os deuses com o sacrifício de um boi. O conto de Ugarit também apresenta as mesmas descrições mitológicas com ligeiras variações (História viva, 2015). Considerando que Abraão viveu na cidade de Ur, que era a cidade dos sumérios, fica evidente que o conto passou por adaptações para construir a versão mitológica de Adão e Eva. Isto demonstra as diferenças mitológicas presentes em diferentes estruturações teológicas e o processo de apropriação cultural dos contos por diferentes correntes religiosas.
Segundo os estudos mais recentes, Gênesis é um livro escrito nos séculos V e VI a.c. (Van Seters, 1998 & Davies 1998) e foi redigido sob quatro fontes, ou estilos literários distintos; Javista, o Elohinista, Deuteronomista e Sacerdotal, cada um contando a mesma história básica, porém, unidas por vários editores diferentes e com ideologias também diferentes (Gooder, 2000).
Segundo os literalistas da bíblia, o dilúvio global ocorreu a 2.348 a.c. (ou seja, 1.652 anos após a criação da Terra, que ocorreu em 4.000 a.c.) Isso significa que o dilúvio ocorreu a 4.363 anos atrás. Segundo Collins da Universidade de Oxford (2001) a pirâmide de Gize foi construída a 3.800 anos a.c. Isso significa que ela foi construída a 4.575 anos atrás.
Quando o dilúvio ocorreu, a pirâmide de Gize já existia á 212 anos. Entretanto, ela não contém marca alguma de qualquer enchente que a cobriu, nem relatos hieroglíficos. Em 2010 o pesquisador chinês Yang Ving Cing afirmou ter encontrado uma estrutura que supostamente seria da arca de Noé, datada então em 4.800 mil anos. Posteriormente a evidência se revelou falsa.
Se cogitarmos que o dilúvio ocorreu á 4.800 anos atrás e a pirâmide de Gize foi construída em 4.575 anos atrás, ou seja, 225 anos após o dilúvio; Como pode ser possível que em somente pouco mais de 220 anos houvesse uma população de escravos com mais de 100 mil homens para construí-las?
Não havia “apenas” 100 mil escravos, mas havia gregos que na mesma época relataram o uso de trabalho escravo e acampamentos dos trabalhadores nas proximidades associados à construção de Gizé. Segundo Verner (2001) o trabalho de construção da pirâmide de Gize foi organizado em uma hierarquia específica, que consistia em 100 mil homens divididos em cinco grupos de 20 mil escolhidos de acordo com as habilidades trabalhistas. Isso nos leva a concluir que se a pirâmide foi construída antes do dilúvio, ela deveria conservar marcas geoquímicas em sua estrutura, o que não ocorre. Se a pirâmide foi construída depois do dilúvio, a contagem de homens para a construção desafia a taxa de repovoamento, afinal, como seria possível mais de 100 mil homens estar presentes no Egito partindo de uma única família que sobreviveu a um dilúvio 300 anos antes?
Segundo o relato, Noé começou a construir a arca com 480 anos, e terminou com 600 anos de idade. Noé viveu até 950 anos e em nenhum momento é relatado no Egito um dilúvio ou mesmo o nome de Noé como protagonista de tal história. Se Noé viveu mais de 350 anos após o dilúvio, ele deveria ter sido citado pelos egípcios após o dilúvio tamanho foi o feito de seu Deus; castigar a humanidade toda com morte e água.
Há duas datas possíveis para o dilúvio; 4.363 ou á 4.800 anos atrás; considerando que tipos básicos devem ter se diversificado em muitas espécies em apenas pouco mais de 4 mil anos, e claro, que a taxa de especiação depende da frequência de reprodução da espécie, animais que demoram para alcançar a idade reprodutiva demorariam ainda mais tempo para dar origem a novas espécies, em geral, vertebrados.
Se usarmos os cães como referência para uma breve estimativa teremos ideia de como o processo funciona. Os baramins de canídeos estão representados na primeira imagem do presente texto. Notamos que o baramin representa todos os canídeos existentes, e sabemos que nem todos os canídeos conseguem hibridizar entre si. Portanto, canídeos e suas 37 espécies existentes atualmente (Sillero-Zubiri, 2009) não suportam um relacionamento baraminológico em apenas 4.363 anos de taxas de especiação. Isto daria uma média de uma espécie nova de Canídeo surgindo a cada 117 anos. Nenhum mamífero têm uma taxa de especiação como esta.
Considerando que canídeos como o lobo atingem a maturidade sexual após dois ou três anos então teríamos no máximo 48 gerações entre uma espécie e outra. Esta data é inconsistente com as evidências fósseis e genéticas.
Usando seleção artificial com raposas, o russo Lysenko, biólogo amigo pessoal de Stalin, conseguiu depois de 35 gerações 78% dos animais se comportando como cães, anatomicamente bastante semelhantes, com uma pelagem malhada em branco e preto. As orelhas caíram, semelhante ao ocorrido com o cão cocker, a ponta da cauda fazia uma curva para cima e o cio da fêmeas ocorria a cada seis meses. A vocalização lembrava a dos cães. Os traços caninos dessas raposas não estavam no objetivo dos pesquisadores, mas acompanharam o processo de seleção artificial que modificou totalmente o comportamento, a anatomia e a fisiologia do animal para uma categoria nova. Mas ainda sim eram apenas raposas domesticadas, e se de fato contemplam uma nova espécie, foi devido á seleção artificial (Dawkins, 2009).
Notamos então que nenhuma dessas alegações baraminológicas faz justiça aos seus pressupostos, e que o tempo de especiação após o dilúvio é escasso para explicar a diversidade de formas de vida fruto de uma “evolução” baraminológica. Imaginemos agora um elefante, cuja maturidade sexual demora entre oito e 12 anos de idade para ser alcançada, ou de uma girafa, cujo tempo é de 3 a 4 anos.
Somando a isto, há o fato de que o relato do dilúvio também é minado com incertezas de datação e em comparação com informações que culturas mais antigas que a do cristianismo. Vimos que o relato de enchentes antecede a origem das religiões judaico-cristãs e de fato, até mesmo culturas posteriores relatam contos de dilúvio com datas muito posteriores a oferecida pelos criacionistas.
Na mitologia hindu, textos como o Satapatha Brahmana mencionam a história de um grande dilúvio em que o Matsya, o avatar do deus Vishnu, adverte o primeiro homem, Manu, sobre a enchente e também aconselha-o a construir um barco gigante. O livro de Satapatha Brahmana foi escrito composto na primeira metade do primeiro milênio a.c. e é um conhecido texto religioso hinduista sânscrito em prosa que descreve os rituais védicos (David, 2004).
Escavações no Iraque, antiga Mesopotâmia, revelaram evidências de inundações localizadas em Shuruppak (atual Tell Fara) e várias outras cidades sumérias. Uma camada de sedimentos fluviais, datada de cerca de 2900 a.c interrompe a continuidade do assentamento que se estende para o norte até a cidade de Kish e que assumiu a hegemonia após o dilúvio. Cerâmicas policromadas do período Jemdet Nasr (3000-2900 a.c) foram descobertas abaixo do estrato da inundação em Shuruppak.
Outros locais, como Ur (cidade dos sumérios, onde Abraão viveu posteriormente), Kish, Uruk (cidade que desenvolveu agricultura e contava com mais de 30 mil pessoas), Lagash e Nínive, também apresentam evidências de inundações. Entretanto, estas evidências vêm de diferentes épocas e períodos. Geologicamente, a inundação de Shuruppak coincide com o surgimento do deserto do Saara e parece ter sido um evento localizado causado pelo represamento do rio Karun através da disseminação de dunas por inundações no Tigre e devido a chuvas fortes em simultâneo na região de Nínive, o que elevou toda a água o rio Eufrates. Em Israel, por exemplo, não existe evidência de uma inundação generalizada. Dadas as semelhanças na história do dilúvio da Mesopotâmia e do relato bíblico, parece que ambas têm uma origem comum (Hendel, 1987). O que demonstra que a grande quantidade de descrição de um relato não implica que ele seja factual, mas que seja um mito compartilhado pelos ancestrais passados oralmente.
Na América do Sul também temos nosso próprio mito diluviano. O povo Tupi foi um dos povos indígenas mais importantes do Brasil. Os estudiosos acreditam que eles se estabeleceram pela primeira vez na floresta amazônica, a cerca de 2.900 anos atrás, eles começaram a se espalhar para o sul e gradualmente ocupando a costa do Atlântico (Ribeiro, 1997).
Alguns autores suspeitam que, nesta trajetória, os tupis tenham enfrentado os tupinambás, que já habitariam o litoral; outros sustentam que apenas se tratava de levas sucessivas do mesmo povo, os posteriores encontrando os anteriores já estabelecidos. Certo é que, nesse processo, as tribos tupis derrotaram as tribos tapuias que já habitavam o litoral brasileiro, expulsando-as, então, para o interior do continente, por volta do ano 1000 (Bueno, 2003).
A mitologia Tupi guarani afirma que durante o tempo conhecido como “Tenonde Yvy” (primeira terra), homens e deuses viviam livremente em abundância e não havia doenças ou dificuldades. Até que um homem chamado Jeupié transgrediu o tabu máximo pela prática do incesto. Ele copulou com a irmã de seu pai. Este ato levou a punição por inundação (mba’e-megua guasu) que destruiu essa primeira terra e produziu à morada celestial. Ñamandú, deus principal do Guarani, decidiu então criar uma segunda terra, agora imperfeita. Pediu a ajuda de Jakairá que espalhou uma névoa na nova terra. Sobreviventes do dilúvio guarani passaram a habitar esta terra, onde há agora a doença, sofrimento e morte. Desde então homens da “nova terra” são chamado Yvy Pyahu estão condenados á eterna busca da terra perfeita perderam, chamando ela de “Yvymara’neỹ” (Terra Sem Mal) (Colman, 1929).
Os Incas surgiram a 2 mil e viveram até aproximadamente 467 anos atrás. Eles dominaram territórios do Chile, Andes, Argentina, Bolívia, Peru e Equador e mantiveram um excelente padrão de desenvolvimento agrícola. Na mitologia Inca, Viracocha destruiu gigantes com uma grande inundação, e duas pessoas repovoaram a Terra: Manco Capac e Mama Ocllo. Eles sobreviveram apenas em cavernas (Gamboa, 2001).
O Lago Titicaca era o lar de um grupo indígena conhecido pelo nome de Auroques ou Urus, há uma lenda local que depois do dilúvio, o lago Titicaca foi onde os primeiros raios do sol surgiram.
Para os kawésqar, ou Alacalufes da Tierra del Fuego (Patagônia), uma grande inundação ocorreu no mundo por punição a um jovem que caçou uma ave para oferecer uma refeição a sua namorada. Porém, o animal era sagrado e não podia ser caçado. Como vingança os deuses trouxeram o mar e matou as pessoas afogadas. No final da história, o jovem e sua namorada são salvos por terem escalado montes elevados e repovoaram toda a terra (Chapman, 2010).
De acordo com uma tradição Taino, no Caribe o deus Yukahua Yukiyu criou uma grande inundação por motivos desconhecidos e só sobreviveu que permaneceu no rio da floresta Yunque (Fray, 1999).
Todos estes relatos são anteriores ou muito posteriores ao relato bíblico e carregam seus deuses, motivos e narrativas específicas. Não existe um relato oficial factual de dilúvio, e nem evidências de que seja um evento global. As pressuposições baraminológicas foram tiradas em cima de pressupostos do mito da criação e pressupostos sobre o relato diluviano. O que notamos então é que todas as premissas criacionistas se consolidam em especulação sobre especulação, sem qualquer argumentação lógica ou empírica. O único substrato que resguarda a baraminologia é uma fé incondicional a um sistema estatístico criado para forjar resultados que contemplem a fé individual, e não que contemple os fatos.
Victor Rossetti
Palavras chave: NetNature, Rossetti, Baraminologia, Homobaramins, Apobaramins, Monobaramins, Polibaramins, Design Inteligente, Criacionismo, Dilúvio, Arca de Noé, Pseudociência.
.

Com toda a certeza você não conhece e não faz a mínima ideia do que seja mutação cumulativa pra afirmar as ‘coisas’ que você afirma sem uma única referencia cientifica a respeito. E veja a ironia, e o tiro no pé, que é o final do seu comentário…”Existia a genética a 400 anos atrás?” – Resposta(?): Não, claro que não existia…mas veja que interessante, esse conhecimento CORROBORA a teoria sintética da evolução!! Ou seja, a genética mostra cabalmente que a evolução ocorre!!
Quando você precisar de um exame de paternidade, você irá aceita-lo ou irá desconsiderar e negar as informações naquele exame? – Então, (acreditando que você irá aceita-lo) esse mesmíssimo conhecimento diz LITERALMENTE que os seres humanos e os grandes símios possuem parentesco.
Reveja seus conceitos e leia livros a respeito antes de emitir opiniões…evita que você passe vergonha! 🙂
E como sempre apanhando