Ao que parece, a relação evolutiva entre procariotas e eucariotas criou vias bioquímicas bastante complexas e diversificadas, especialmente no que diz respeito a fotossíntese em cianobactérias e as algas. Durante muito tempo, a bioquímica da fotossíntese foi considerada muito complexa e muito semelhante em detalhes para ter surgido duas vezes de modo independentemente. Portanto, concluiu-se que todos os organismos fotossintetizantes são historicamente relacionados e, por extensão, cianobactérias evoluíram para eucariotas fotossintéticos. Este eucarioto ancestral era uma alga vermelha porque seus pigmentos e antenas moleculares mais se assemelham as encontradas em cianobactérias, que também não têm flagelos e corpos basais (Dougherty et al, 1960).
Archamoeba
A fotossintese é muito semelhante entre cianobactérias e eucariotos fotossintéticos. Isso porque os plastídios das células vegetais e algas são derivados. Surgiram a partir de um simbionte com cianobactérias. Com o aumento da aceitação da origem dos plastídios a partir de cianobactérias, as ligações entre cianobactérias e o núcleo podem explicar a origem dos eucariotos (eucariogênese) (Keeling, 1998).
Uma alternativa para essa proposta surgiu com Carl Woese e seus colegas, que descobriram uma divisão inesperada em procariontes, em 1977. O grupo de Woese mostrou que procariontes são compostos de dois grupos relacionados, dois quais ele chamou de Eubacteria e Archaebacteria, hoje sinônimo de Bactérias e Archaea. Os Archaebacteriologistas começaram a encontrar as ligações moleculares entre Archaea e Eucariotas (Huet et al, 1983). Mais adiante veremos críticas quanto a esta classificação.
Eles concluíram que Archaeas compartilham um ancestral comum recente com eucariontes (Iwabe et al, 1989). Cada uma dessas semelhanças compartilhadas esclarece a transição entre procariotos/eucariotos em um pequeno caminho, mostrando que as características moleculares previamente consideradas ”eucarióticas”, estavam presentes anteriormente a origem das células nucleadas (do grego ευ que significa “bom, perfeito”; e κάρυον, que significa núcleo).
No entanto, este compartilhamento de características liga Archaeas com todos os eucariotas, diferente da hipótese que liga cianobactérias a algas, e nenhum grupo de eucariotas assemelha-se especificamente a Archaeas. Então, qual foi a natureza do primeiro eucarioto?
Para Margulis, e a hipótese eocítica, isso ocorre porque eucariotos são organismos quiméricos, mosaicos com caraterísticas de Archaeas e particularidades próprias fruto da endossimbiose. Certas células eucarióticas começaram a atrair a atenção dos biólogos ultimamente. Se a mitocôndria, assim como os plastídios se originou em um evento endossimbiótico, é possível que algumas linhagens eucariotas se tornassem amitocondriadas (sem mitocôndrias) ao divergir antes desse evento endossimbiótico (Keeling, 1998).
Esta foi a primeira proposta sugerida ao grupo chamado de hipermastigotos, que compreende especificamente o grupo de Parabasalia (Stewart & Mattox, 1980) que foi estudado, ampliado e refinado por Cavalier-Smith em 1983. Ele propôs o grupo Archezoa que contém os descendentes de antigos eucariotas pré-mitocondriais, e dividiu tal grupo em: Metamonadas, Parabasalia, Microsporidia e posteriormente, os Archamoebae. Cada um destes grupos é composto por seres anaeróbicos, células amitocondriadas que são morfologicamente muito simples. Além de não têm mitocôndrias, falta em alguns membros os hidrogenossomas, peroxissomos ou microcorpos como os dictiossomos de Golgi. Em alguns grupos, nem mesmo há flagelos.
Ribossomos eucarióticos tem seu RNA caraterizado pelo tamanha, definido como 80S, enquanto que em Archezoa e procariontes são 70S. Do mesmo modo, as moléculas de RNAr eucarióticos são de 18S e 28S maior parte, em contraste com os rRNAs menores encontrados em procariotas e Archezoa (Keeling, 1998).
Filogenias moleculares foram produzidas para mostrar o relacionamento desses grupos, geralmente usando pequenos genes de RNAr (subunidades de microsporídio e Diplomonadida/Metamonada), genes de Parabasalia de modo comparativo (Vossbrinck et al, 1987), a expressão gênica de EF-1a, EF-2, subunidades de RNA-polimerase, isoleucil-RNAt-sintetase, a grande subunidade de rRNA. Todos corroboraram as relações evolutivas e posição dos grupos com suas respectivas ramificações (Brown & Doolittle, 1995).
Até então, a hipótese mais aceita é que Archezoa são anteriores à origem da mitocôndria, então eles devem ramificar mais cedo do que eucariotas contendo mitocôndria em árvores filogenéticas. Entretanto, surgiram evidências de que vários destes organismos contêm um resíduo genético de mitocôndrias.
As evidências dizem que Archezoa está perdendo rapidamente seus componentes adquiridos em uma endossimbiose mitocondrial que pode ter ocorrido antes da evolução de qualquer das linhagens eucarióticas atualmente conhecida. As evidências estão em cada um dos grupos de Archezoa
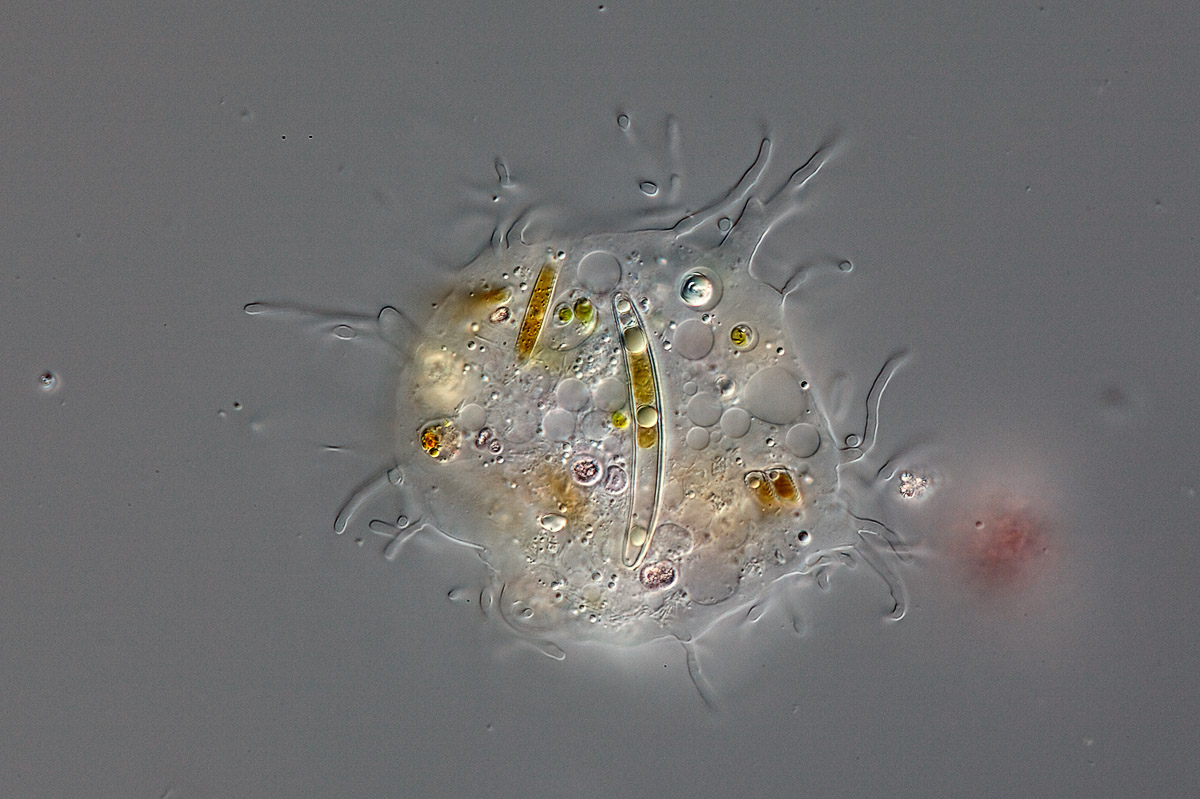
Archamoebae é um grupo que compartilha características que unificam todas as amebas sem mitocôndria. Formam um grupo bem diversificado habitando uma grande variedade de ambientes. A diversidade da Archamoebae tem sido motivo para se duvidar da unidade filogenética do grupo (Cavalier-Smith, 1987), embora, ao mesmo tempo sua simplicidade incomparável com os membros também levanta a sugestão de que de todos Archezoa (e claro, a Archamoebae) são muito primitivos (Cavalier-Smith, 1991). Isso significa que carregam características muito antigas, de linhagens antigas embora sejam grupos presentes ainda hoje. (lembrando que características primitivas não indica que um o animal seja um fóssil vivo. Pode ser simplesmente uma Simplesiomorfia).
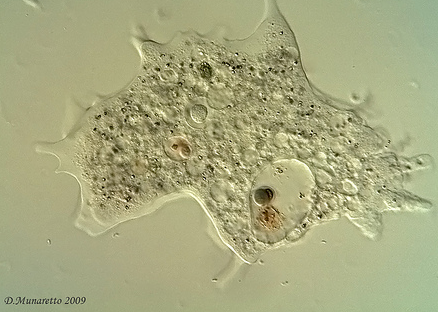
Amoeba Pelomyxa (Archamoeba). Clique para ampliar
Uma evidência de que a Archamoeba pode ter se tornado secundariamente amitocondriada veio do patógeno entérico Entamoeba histolytica. Quando o gene da subunidade pequena rRNA de Entamoeba foi caracterizado, percebeu-se que esta espécie ramificou-se mais tarde do que alguns protistas contendo mitocôndrias (Herolobosea e Euglenozoa), que sugeriram do ancestral de Entamoeba que tinha de fato mitocôndria e a conserva até os dias atuais (Sogin, 1989).
Desde então, a pequena subunidade de rRNA de Phreatamoeba balamuthi e a grande subunidade de rRNA de Pelomyxa também foram sequenciadas comparando com os eucariotas que contém mitocôndrias (Hinkl et al, 1994 e Morin & Mignot, 1995) demonstrando que tal grupo surgiu a partir de ancestrais contendo mitocôndrias. Na pequena árvore da subunidade rRNA, Entamoeba e Phreatamoeba se ramificar em conjunto (Cavalier-Smith & Chao, 1997) e às vezes não (Hinkl et al, 1994). Isso suscita dúvidas sobre a validade de Archamoebae como uma linhagem evolutiva, exatamente pela posição em que esta, mitocondriado ou primitivamente amitocondrial.
O genoma de Entamoeba carrega relíquias moleculares do simbionte mitocondrial. A maioria das muitas centenas de genes codificadores de proteínas mitocondriais são codificadas no núcleo e orientados para a organela depois de serem traduzidos no citoplasma. Estes genes foram transferidos a partir do genoma do simbionte para o núcleo, mas são reconhecíveis hoje por causa deste direcionamento e porque os próprios genes homólogos se assemelham a partir do tipo de bactéria a partir do qual as mitocôndria evoluíram, uma α-proteobacteria (Yang e tal, 1985).
Encontrar tais genes no núcleo de um organismo mostra que os antepassados de que continha uma linhagem mitocôndria.
Isso revelou um avanço para testar o grupo Archezoa como um todo. Clark e Roger (1996) encontraram dois genes de origem mitocondrial em Entamoeba, um para a piridina-transhidrogenase e outro para uma chaperonina 60-kDa (chamada de cpn60). Em análises filogenéticas de cpn60, a proteína de Entamoeba ficou fortemente relacionada com os homólogos da mitocôndria de outros eucariotas, que por sua vez foram relacionadas com as proteínas de cpn60 de α-proteobacteria.
A explicação mais simples é que Entamoeba evoluiu de ancestrais mitocondriais, (Clark & Roger, 1996) porque a chaperonina deste grupo não só está de acordo com as expectativas filogenéticas de uma proteína derivada da mitocôndria, mas também é especificamente relacionada com homólogos eucarióticos que são direcionados diretamente para as mitocôndrias.
Outro grupo que evidência essa relação entre Archezoa e mitocondrias é o Parabasalia. Eles contêm uma organela metabólica com membrana dupla chamada hidrogenossoma cuja origem tem sido a fonte de grande debate.
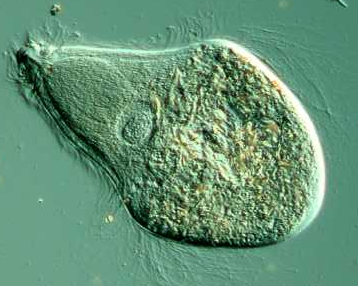
Parabasalia (Trichonympha)
Os hidrogenossomas de Parabasalia não se assemelham as mitocôndrias em sua morfologia (com exceções), eles não contém um genoma e, ao contrário da fosforilação oxidativa na mitocôndria, a energia é liberada no hidrogenossoma a partir da conversão de piruvato ou malato
em acetato, dióxido de carbono e hidrogênio.
Ainda sim, houve suspeitas de que hidrogenossomas de Parabasalia podem partilhar ancestralidade comum com mitocôndrias. Isso porque, hidrogenossomas não estão restritos a este grupo, mas são encontrados em membros isolados de várias linhagens eucarióticas independentes (Percolozoa, Ciliados e Fungos Chytrid), e estes organismos não têm invariavelmente mitocôndrias. Alguns desses hidrogenossomas se assemelham morfologicamente a mitocôndrias (Finlay & Fenchel, 1989).
A sugestão de que as mitocôndrias podem ter se transformado em hidrogenossomas nestes organismos, poderia retirar Parabasalia dos Archezoa (Cavalier-Smith & 1987). No entanto, em alguns destes organismos a hidrogenossoma também tem sido defendido por ter evoluído a partir de peroxissomas (Marvin-Sikkema, 1993) e tendo em conta as diferenças entre hidrogenossomas e mitocôndrias, sua exclusão mútua não é, por si só, evidência suficiente de que eles são de ancestrais (Muller, 1993).
Genes de chaperonina (cpn10, cpn60, e cpn70) foram encontrados no genoma do Trichomonas e usados para tentar resolver esta questão. Em cada caso, estes genes foram especificamente relacionados com homólogos mitocondriais (Horner et al, 1996 e Roger et al, 1996).
Em testes com anti-corpos, cpn60 e cpn70 de T. vaginalis reagem especificamente com as hidrogenossomas purificados (Bui et al, 1996) e anticorpos de cpn60 bacterianos foram relacionados com hidrogenossoma (Bozner, 1997). Estas chaperoninas são aparentemente localizadas nos hidrogenossomas e derivados a partir do mesmo endossimbionte como a mitocôndria, criando um forte argumento de que o hidrogenossoma de Parabasalia e das mitocôndrias descendem de um ancestral comum.
Nos ciliados e fungos, onde os hidrogenossomas evoluíram a partir de mitocôndrias, eles vieram de uma organela altamente especializada porque mitocôndrias estavam bem desenvolvidas nos ancestrais desses grupos. Mas isso não é necessariamente o caso de Parabasalia onde mitocôndrias não são encontradas em qualquer linhagem anterior a Parabasalia. Por isso, tudo o que pode ser dito é que o hidrogenossoma evoluiu a partir do mesmo simbionte.
Se os Parabasalia ancestrais tinham uma mitocôndria, que em seguida, formou os hidrogenossomas, pode ter ocorrido com ciliados e fungos. Este processo implica a perda das enzimas metabólicas essenciais encontradas em mitocôndria e o recrutamento das enzimas não-mitocondriais, como hidrogenases e de piruvato: ferredoxina oxidorredutase de alguma outra fonte.
Se hidrogenossomas vieram de um simbionte não-especializado, seguindo uma trajetória evolutiva diferente do que fez a mitocôndria, e isto reflete a probabilidade de que o simbionte teve uma maior diversidade metabólica do que as atuais mitochondrias (Muller, 1997).
Microsporidia são parasitas intracelulares obrigatórios. Fungos podem sobreviver como esporos com um duplo revestimento de quitina resistente. No interior do citoplasma há uma estrutura enrolada conhecida como o tubo polar. Esta organela parece ser um híbrido entre um “arpão” e uma “agulha hipodérmica”. Quando um esporo encontra um hospedeiro susceptível, ele rapidamente everte o tubo polar, que, em seguida penetra na membrana do hospedeiro. O citoplasma é espremido através do tubo polar e é injetado diretamente no citoplasma do hospedeiro onde vive como uma ameba, dividindo-se e produzindo mais esporos (Keeling, 1998).
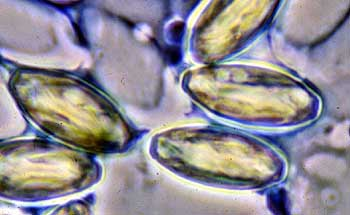
Microsporidia
Microsporidia são parasitas talentosos. Tem os menores genomas nucleares conhecidos, menor até mesmo que os de bactérias (Biderre et al, 1995).
Os ribossomas de Microsporidia assemelham-se com o dos procariotas em que a sequência homóloga à molécula de rRNA 5.8S está ligada de forma covalente com o RNAr 23S; em outros eucariotas este RNAr é uma molécula separada (Vossbrinck& Woese, 1986). A fusão de 5.8S-23S é única para microsporídeos e procariotas e tem sido citada por suportar a ideia de que esses fungos são os mais primitivos de todos eucariotos (Siddall et al, 1993).
No entanto, o gene 5.8S está localizado na porção de cima do gene da subunidade grande no operon de DNAr em eucariotas. Quando esse operon é transcrito estes dois rRNAs são clivados em locais específicos de transformação. Sequências de rRNA de microsporídio são estranhas em geral, contêm muitas exclusões. Se uma dessas supressões afetou um único local de processamento em um ancestral de Microsporidia, ele poderia ter conduzido à reforma dos rRNA 5.8S-23S fundidos (Cavalier-Smith, 1993).
Todas essas características podem ser evidências de que Microsporidia são descendentes antigos de eucariontes primitivos altamente adaptados, parasitas, o que é característico do grupo.
Estudos sobre a natureza de sua história evolutiva ainda são iniciais e estão cheios de surpresas. As evidências atuais estão sugerindo que Microsporidia realmente evoluiu de forma “soberana” entre os eucariotas (dos galhos dos animais e plantas/fungos) e podem compartilhar um ancestral próximo com fungos.
A evidência mais forte suporta uma relação geral entre Microsporidia e os taxa soberanos. Isso porque as proteínas EF-1a de microsporídeos, animais, fungos e contêm todas uma inserção que é única para estes taxa (Kamaishi et al, 1996) Da mesma forma, a di-hidrofolato-redutase e timidilato-sintase são duas enzimas separadas em microsporídios, animais e fungos, mas são fundidas em plantas e outro protistas (Vivare`s et al, 1996). Uma relação específica com fungos foi proposta pela primeira vez com base em paralelos encontrados entre o ciclo meiótico incomum de Microsporidia. Certos grupos de fungos (Flegel & Pasharawipas, 1995) também são apoiados pela filogenia de α e β-tubulinas (Germot et al, 1997).
Em Microsporidia foram descobertos genes mitocondriais (cpn70), nos genomas de Nosema locustae (Germot et al, 1997) e Vairimorpha necatrix (Hirt et al, 1997). Em árvores filogenéticas estas sequências se ramificam de forma convincente com os homólogos mitocondriais, minando o argumento de que eles são descendentes primitivos de eucariotas amitocondriados. Os genes cpn70 de microsporídio também se ramificam, de maneira fraca, com os genes mitocondriais de fungos (Germot et al, 1997 & Hirt et al, 1997)
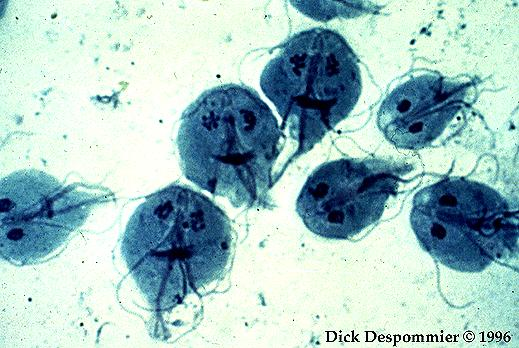
Metamonada (G. lamblia)
O outro grupo que também apresenta um caso interessante é chamado de Metamonadas, que parece abrigar relíquias mitocondriais. Dentro dos Metamonadas há um sub-grupo denominado Diplomonadas. Este, provavelmente é o mais altamente derivado dos Metamonadas, e mal representa o estado ancestral do grupo. Há Metamonadas flagelados, com um único núcleo e quatro cinetossomos, um dos quais é muitas vezes associado ao flagelo com uma organela que auxilia na alimentação, o citostoma. A maioria dos Diplomonadas são formados essencialmente por duas células fundidas em simetria axial. Existem dois núcleos associados com quatro cinetossomos cada, e em certas espécies heterotróficas há também dois citostomas simétricos.
Supostamente, Metamonadas nunca abrigaram uma mitocôndria e portanto não contêm organelas comparáveis aos hidrogenossomas dos Parabasalia, e, ao contrário de Microsporidia, suas seqüências moleculares não são tão incomuns a tal ponto se destacarem-se como candidatos a árvores filogenéticas.
Mas os genes de transcrição e de tradução de outros genes colocam-os consistentemente na base da filogenia de eucariotas (Klenk et al, 1995).
Um estudo revelou que Giardia lamblia (Diplomonada) contém proteínas que reagem a anticorpos mitocondriais cpn60 (Soltys & Gupta, 1994).
Uma possibilidade é que esta chaperonina seja derivada da mitocôndria, mas assumiu um papel citosólico. Isto contrasta com outras proteínas chaperonina em eucariotas amitocondriais: como as chaperoninas de Trichomonas que são voltadas para os hidrogenossoma (Bui et al, 1996) e ambos os genes de chaperonina de Vairimorpha e Nosema que provavelmente dirigem proteínas para alguns compartimentos celulares ainda não identificados (Germot et al, 1997 & Hirt et al, 1997).
O processo pelo qual uma proteína de origem organelar assume um papel citosólico é chamado de “realocamento”. Estas proteínas perderam todos os vínculos funcionais com a organela, mas sua história evolutiva é revelada em sua posição filogenética entre as bactérias.
Isso ocorre, por exemplo, com a 3-fosfoglicerato-quinase em plantas, porque a proteína citosólica está mais intimamente relacionada com os homólogos de plastídios e bactérias do que com outros genes citosólicos de eucariotos.
Há também razões para suspeitar que certas proteínas glicolíticas em eucariotas podem ser derivada a partir de um simbionte mitocondrial. Estas proteínas estão mais relacionadas com homólogos de proteobactérias (os parentes mais próximos da mitocôndria) do que homólogos de arqueobactérias (os parentes mais próximos da linhagem nuclear citosólica que veremos mais adiante) (Keeling, 1998).
Esta relação foi vista em 3-gliceraldeído-fosfato desidrogenase (GAPDH) (Henze et al, 1995) e triosefosfato-isomerase (TPI) (Keeling & Doolittle, 1997). Nesses casos, os genes de Diplomonadas são conhecidos e não diferem notavelmente dos de outros eucariotas. Nem GAPDH e tão pouco o TPI são totalmente inequívocos. A filogenia de GAPDH é complexa, com inúmeras famílias de genes parálogos distribuídos em um padrão cuja interpretação ainda não está clara (Henze et al, 1995) e em TPI, ainda não há informação filogeneticamente suficiente e útil para discriminar entre uma origem especificamente a partir de uma α-proteobacterial a em eucarióta ou simplesmente o de proteobactérias em geral (Keeling & Doolittle, 1997)
As proteínas eucarióticas que parecem estar mais próximos de homólogos de proteobacterias do que de Arqueobactérias têm sido geralmente interpretada como o resultado de transferências laterais ou mesmo um antigo evento de fusão celular, o quimerismo (Rosenthal et al, 1997).
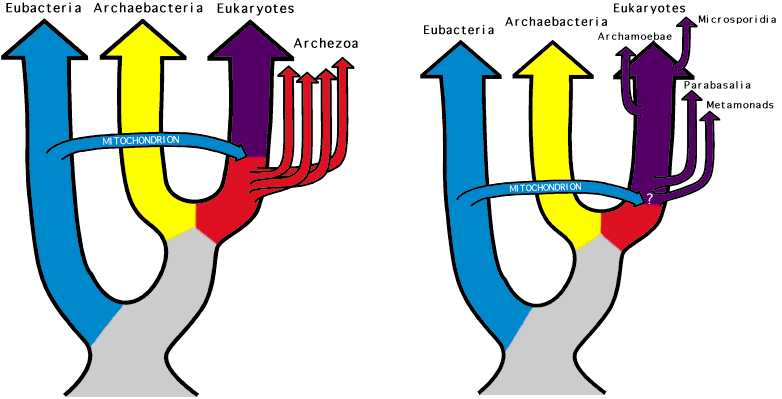
Esquerda: O grupo archezoa como originalmente foi concebido Os Eucariotos com mitocôndrias (roxo), Archezoa (vermelho), Eubacteria (azl) e Archaebacteria (Amarelas). O evento de simbiose que levou a incorporação da mitocôndria foi proposto como tendo ocorrido após a divergência de, pelo menos, quatro linhagens de outros eucariotas (Metamonadas, Microsporidia, Parabasalia, e Archamoebae). Supostamente, esses grupos se separaram antes dá origem dos eucariotas com mitocondrias. Direita: Hoje, o grupo Archezoa. O esquema de cores é como na figura anterior, mas Archamoebae e Microsporidia são mostrados com uma ramificação muito posterior do que a versão anterior. Ela põem Parabasalia e Metamonads na ramificação entre os eucariotas, mas após a simbiose mitocondrial. Segundo as evidências.
Infelizmente, para muitos destes genes ainda não temos informações suficientes sobre as sequências ou amostragem do táxon para tirar conclusões mais precisas quanto à sua origem.
Isto significa que, mesmo que os genes de TPI ou GAPDH sejam derivadas do simbionte mitocondrial, a sua presença não é evidência para a presença da organela. Então para Metamonadas ainda fica uma pequena incerteza.
Isso no faz concluir que o objetivo pela qual o grupo Archezoa foi criado uniu eucariotas primitivamente a-mitocondriados (Parabasalia, Microsporidia, ou Archamoebae), e crescentes dúvidas para os Metamonadas persistem.
No entanto, ja se propõem que Archezoa seja explicitamente uma hipótese filogenética
porque são muito provavelmente descendentes dos primeiros eucariontes que tiveram mitocôndria (Keeling, 1998). E talvez a coisa fique mais clara quando notamos a principal característica que diferencia uma célula procariota de uma eucariota; o eukarion, ou seja, o núcleo.
Por ser a principal característica de célula eucariótica, a origem do núcleo tem sido alvo de muitas especulações. Existem quatro teorias propostas para explicar a existência do núcleo; todas aguardando mais dados (Brinkmann & Martin, 1996).
A teoria mais conhecida vem do modelo sintrófico e propõe que uma relação simbiótica entre as Archaea e as Bactéria se iniciou durante a eucariogênese. Acredita-se que a simbiose se originou quando Archaea primitivas, similares às atuais Archaea metanogênicas invadiram e passaram a viver dentro de bactérias similares às atuais Mixobactérias, formando assim um núcleo primordial. Esta teoria é semelhante a origem da mitocôndria eucariótica a partir das α-proteobacteria e do cloroplasto.
A origem do núcleo entre as Archaea é suportado por observações de que este grupo e os eucariotas possuem genes similares para proteínas importantes, incluindo as histonas. As observações mostram mixobactérias como organismos móveis, que podem formar complexos multicelulares e que possuem quinases e proteínas G similares aos Eucariotos, suportando assim uma origem bacteriana da célula eucariótica (Lopez-Garcia & Moreira, 2006).
O segundo modelo propõe que células proto-eucarióticas evoluíram a partir de bactérias sem estágios endossimbióticos. Este modelo é baseado na existência das bactérias do filo Planctomycetos, que possuem uma estrutura nuclear com poros primitivos e outras estruturas membranares compartimentadas (Fuerst, 2005). Um modelo similar propõem que uma célula semelhante à eucariótica, o cronócito, evoluiu primeiramente, tendo depois fagocitado membros das Archaea e Bacteria, gerando assim o núcleo e a célula eucariótica (Hartman & Fedorov, 2002).
O modelo mais controverso é o da eucariogênese viral, que propõe que o núcleo composto de membranas, assim como outras estruturas eucarióticas, originaram-se a partir da infecção de vírus. A sugestão é suportada por similaridades entre eucariotas e vírus: fitas lineares de DNA e fortes ligações a proteicas (analogia entre histonas e envelope viral). Uma versão da proposta sugere que o núcleo evoluiu ao mesmo tempo em que a fagocitose, formando um predador celular primitivo (Bell, 2001). Outra variante propõe que os eucariotas são originários de Archaea primitivos, infectados com Poxvirus, e baseia-se nas semelhanças entre a DNA-polimerase de modernos poxvirus e eucariotas (Takemura, 2001 & Villarreal & DeFilippis, 2006). Ainda é sugerido que a evolução do sexo possa estar ligada à hipótese da eucariogênese viral (Bell, 2006).
Finalmente, uma proposta recente sugere que variantes tradicionais da teoria da endossimbiose são insuficientemente robustas para explicar a origem do núcleo eucariótico.
Este modelo, denominado “hipótese exo-membranar”, sugere que o núcleo se originou de uma única célula ancestral que formou uma segunda membrana celular externa; a membrana interior que envolvia a célula original tornaria-se a membrana nuclear, formando poros mais complexos ao longo do tempo, permitindo a passagem de componentes celulares sintetizados internamente como as subunidades ribossomais.
A origem do Núcleo Eucariótico
Acredita-se que o ancestral comum de todos os eucariotas surgiu pela fusão genômica de dois ou mais diferentes procariotos, gerando “quimeras” via simbiogênese (Golding & Gupta, 1995)
A quimera originou-se quando uma Archaebacteria (um termoacidófilo) e uma eubacteria móvel surgiu sob pressão seletiva; advinda do oxigênio e escassez tanto de compostos de carbono e receptores de elétrons. O núcleo celular evoluíu desse quimerismo.
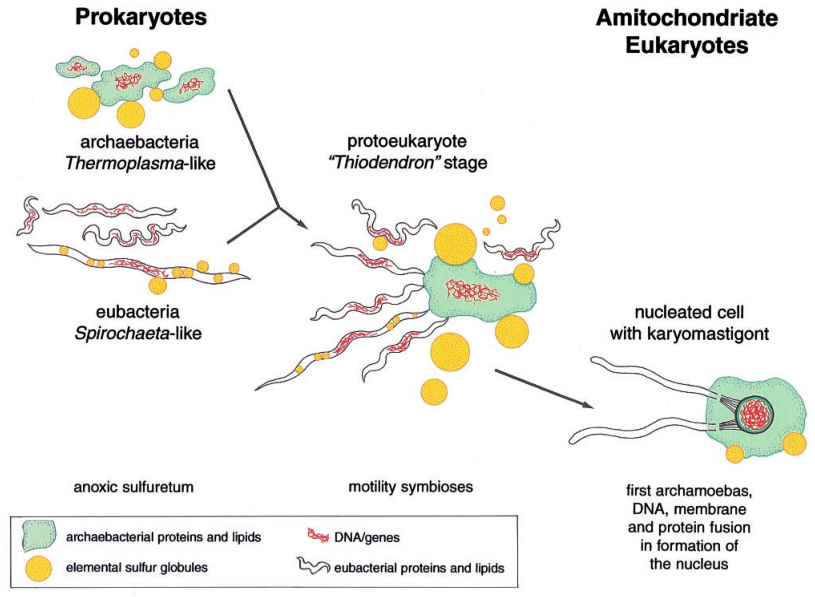
Origem do primeiro eucarioto quimérico com cariomastigonte (karyomastigonte) a partir de um consórcio com uma bactéria móvel do enxofre. Clique para ampliar
Se o mais antigo descendente desta fusão estivesse vivo ainda hoje, seria reconhecido como um protista a-mitocondriado, como vimos acima. Esse paradigma hoje se consolida devido a sua consistência simultânea no cenário evolutivo entre as áreas de ciências: biologia celular, biologia do desenvolvimento, ecologia, genética, microbiologia, evolução molecular, paleontologia, protistologia (Chapman et al, 2000).
A proposta do núcleo como resultado de um processo de fusão vem de Lynn Margulis (Margulis et al, 2000) e em sua análise, ela toma partido de outro ponto vista ligeiramente diferente ao que é proposto inicialmente por Carl Woese. Seu modelo de abordagem se pauta em apenas dois grupos biológicos (Bacteria e Eucarioto) e não no sistema de 3 Domínios biológicos (Archaea, Eubacteria e Eukarya) (Woese et al, 1990).
Sua crítica quanto a este modelo considera que existe contradições na forma de classificação de microrganismos desde 1820. Scamardella (1999) observou que o sistema não-morfológico de classificação de Woese desconsidera simbioses.
Embora alguns procariotas certas bactérias Gram-positivas são intermediárias entre arqueobactérias e eubactérias (Grupta, 1998) mas não há organismos intermediários entre procariotas e eucariotas, embora certos grupos de Archaea hoje preencham lacunas importantes entre esses dois grupos celulares. Entretanto, via de regra, esses fatos tornam os RNAs 16S e outras taxonomias não-morfológicas de Woese inadequadas por serem incompletos.
Quando Woese (1998) declara que há três, e não dois, agrupamentos filogenéticos principais de organismos neste planeta, ele afirma que Archaebacteria e Eubacteria são mais próximos um do outro do que com os eucariotas. Desta forma, ele nega a motilidade intracelular e o núcleo mitótico. Ele minimiza estes e outros dados biológicos celulares, histórias de vida sexuais, incluindo a fusão cíclica celular, correlação com registro fóssil (Margulis, 1996) e as comparações moleculares baseadas em proteínas (Gupta, 1998 e 1999).
Para Margulis, a suposição desinformada de Woese (e de outros biólogos moleculares) de que a hereditariedade reside somente em genes nucleares é contrariada pela literatura científica do desenvolvimento embrionário, citológico e hereditariedade citoplasmática (Sapp, 1999). Os sistemas de motilidade e alimentação com base na tubulina-actina e fusão celular sexual facilitam a a incorporação viável e frequente de ácidos nucleicos heterólogos. Muitos eucariotas e procariotas naturalmente adquirem genes em seus genomas em um único evento fagocítico sem ingerir regularmente células inteiras. Isso invalida qualquer medida isolada, incluindo sequências de genes de RNA ribossomal.
Como quimeras, eucariotas evoluíram por integração de mais do que um único genoma procariótico. Porque procariontes não são diretamente comparáveis com os eucariotas simbioticamente gerados. O mais correto seria rejeitar a interpretação de três domínios de Woese e ver os primeiros eucariotas anaeróbios derivados de ambas linhagens procarióticas (Margulis et al, 2000).
As enzimas da síntese de proteínas em eucariotas vêm principalmente de arqueobactérias enquanto que no sistema de microtúbulos (motilidade e os seus centros de organização), muitos de proteínas solúveis provenientes de eubactérias (Margulis, 1996).
Estudo de sequências de proteínas conservadas (um conjunto de dados muito maior do que a usada por Woese) concluiu (Gupta, 1998) que todas as células eucarióticas, incluindo as a-mitocondriadas e a-plastídicas receberam grandes contribuições genéticas em seu genoma nuclear. Genes provenientes de arqueobactéria (muito provavelmente da Eocite [hoje chamada de Crenarchaeota], ou seja, um termoacidófilo) e de bactérias Gram-negativas. A célula eucarióta ancestral não descendente diretamente de arqueobactérias, mas em vez disso foi uma quimera formada pela fusão e integração de genomas desses dois grupos.
Nesse estudo, o autor, Gupta, afirmar que a contribuição eubacterial sugere que arqueobactérias são um grupo polifilético e são parentes próximos de bactérias Gram-positivas. Isso quer dizer que o grupo que não inclui o ancestral comum de todos os indivíduos, ou ainda, é a reunião de dois ou mais grupos monofiléticos (de origem única). Seus integrantes possuem vários ancestrais comuns, um em cada grupo.
Para Margulis, as sequências archaebacteriais seguem a tendência proposta por Searcy (Searcy, 1992), a partir de um procarioto eócito termoacidófilo. Este antepassado archaebacterial viveu em águas quentes, ácidas, profundas e esporadicamente sulfurosas, usando tanto o enxofre (H2S) ou o oxigênio como receptor de elétrons terminal, como faz seus descendentes. A antiga archaebacteria sobreviveu a condições ambientais de ácido/hidrólise usando um sistema semelhante ao nucleosome-histona envolvendo seu DNA com uma camada proteica (Searcy, 1992), sintetizando proteínas com a actina (ou análogos) (Searcy et al, 1980). A archaebacteria sem paredes foi então, um grupo pleiomórfico. Isso quer dizer tinham a capacidade alterar a sua forma ou tamanho em resposta às condições ambientais.
Outros membros do grupo poderiam também estar ligado a eucariogênese e a gênese do núcleo celular. Sendo ele anaeróbio obrigatório, acaba utilizando condições altamente redutoras, como o enxofre para a redução do sulfato a sulfeto de hidrogênio. A degradação de carboidratos (amido, açúcares, tais como celobiose) e a oxidação do sulfureto a enxofre elementar em eubactérias são gerados como produtos da fermentação rica em carbono e são aceptores de elétrons em arqueobactéria.
Outro modelo que pode explicar a origem do núcleo vem de Thiodendron.
Refere-se a um conjunto de bactérias existentes que dá suporte a ideia de motilidade e sintrofia e simbiose usando o enxofre em arqueobactérias-eubactérias. Esses “parceiros” se uniram para formar o antecessor quimérico de Arqueoprotistas. O núcleo delimitado por uma membrana, é a manifestação morfológica do sistema genético quimérico que evoluiu partir de um consórcio como o Thiodendron.
Cada fenômeno sugerido (a partir de bactérias de vida livre a uma associação integrada), tem análogos naturais existentes. O estudo da microbiota marinha revelou consórcios bacterianos relevantes em mais de seis locais geograficamente separados. As amostras foram tomadas a partir de regiões anóxicas (Dubinina et al, 1993). As análises laboratoriais mostram que é necessário abolir o gênero Thiodendron, porque é ele é um sintrófico de enxofre.
Uma associação ectossimbiótica estável de dois tipos de bactérias cresce como uma associação anaeróbica entre 4 e 32 °C em valores de pH e salinidade marinhas. Amido, celobiose, e outros hidratos de carbono (não de celulose, aminoácidos, ácidos orgânicos, ou álcool) suplementados por fixação heterotrófica de CO2 fornece o carbono. Thiodendron aparece como colônias branco-azuladas esféricas gelatinosas, concêntricas na estrutura interna formando uma matriz viscosa produzida por essa associação. O parceiro dominante é invariavelmente uma cepa distinta de espiroquetas (Margulis et al, 2000).
O outro parceiro é um vibrioide, morfologicamente estável, do gênero Desulfobacter sp. Ele necessita de carbono orgânico, principalmente acetato, advindo da degradação de carboidratos das Spirochetas. Uma espiroqueta semelhante a Escherichia coli fermentadora de ácido fórmico gera energia e alimentos. Células de Desulfobacter sp. reduzem tanto sulfato e enxofre em sulfureto e estão sempre presentes formando essa associação natural, mas em muito menor abundância do que as outras espiroquetas.
O Thermoplasma, produz sulfureto e libera pequenas quantidades de oxigênio para manter baixo potencial redox na sua associação com espiroquetas. Os antecessores sintróficos quiméricos eram metabolicamente análogos ao Thiodendron onde Desulfobacter reduzia enxofre e sulfato para produzir sulfureto em níveis que permitam as espiroquetas crescer.
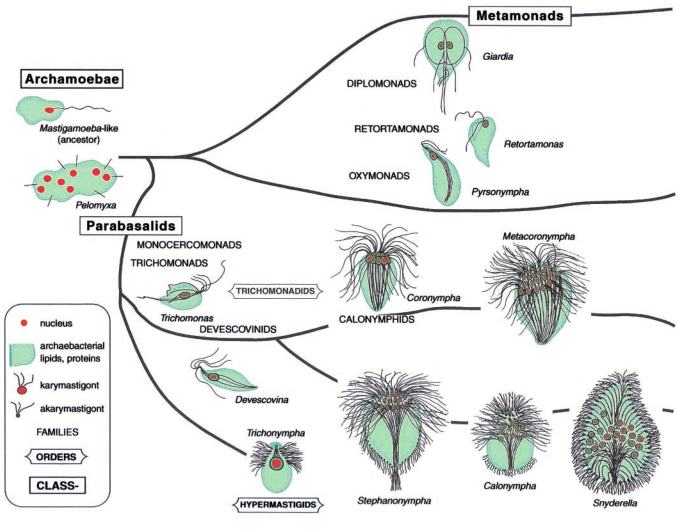
Filogenia de eucariotas quiméricos que podem ser primitivamente a-mitocondriadas.
O DNA da arqueobactéria Thermoplasma é semelhante e permanentemente recombinado com o de eubactérias. Existe um precedente para a sugestão de que hipertrofias na membrana em torno de DNA possam ter formado uma vesícula estável em alguns procariotas, como ocorre na membrana nucleóide das eubacterias Gemmata obscuriglobus (Fuerst & Webb, 1991).
O parceiro natante anexado a elas ofereceu o precursor do sistema mitótico através dos microtúbulos que pertenciam a gêneros quase onipresentes nesse consórcio de espiroquetas ou dos blocos de cntruçao de tubulinas citoplasmática presentes em uma espécie do gênero Hollandina (Margulis, 1993).
A proposta mais enriquecedora a respeito da origem do núcleo, e que favorece a eucariogênese vem de um grupo chamado de Cariomastigonte.
Como vimos, Parabasalia e Oxymonadida (um Metamonada) não possuem mitocôndrias. E esses grupos hoje estão unidos por terem flagelos ou corpos basais em característicos grupos de quatro, muitas vezes associados com o núcleo celular, formando uma estrutura denominada cariomastigonte.
Esse termo foi criado por Janicki (Janicki, 1915) para se referir a um sistema organelar conspícuo encontrado em certos protistas: um axonema mastigonte microtubular (estrutura contrátil dos flagelos e cílios, formada por nove pares de microtúbulos que se prolongam do corpo basal e um par central, além de proteínas associadas) que é sustentado por um cinetossomo ligado por um conector nuclear (rizoplasto). A necessidade do termo veio quando Janicki estudava a motilidade em simbiontes de Trichomonadas presentes no intestino de cupins, onde cariomastigotas dominam as células.
Quando os cinetossomos, conectores nucleares e outros componentes estavam presentes e o núcleo ausente na sua posição previsível, Janicki chamou o sistema organelar de akaryomastigonte.
Na família Calonymphidae, onde estão as tricomonas inteiramente multinucleadas, muitos cariomastigontes e acariomastigontes estão presentes na mesma célula, por exemplo, Calonympha grassii (Kirby & Margulis, 1994).
O cariomastigontes tem características ancestrais de eucariotas, e está presente em muito “cedo” na ramificação dos protistas (Dacks & Redfield, 1998 & Edgcomb et al, 1998). Archaeprotistas formam um grande táxon são unicelulares heterotróficos que habitam ambientes anóxicos. Todos possuem mitocôndrias.
Pelo menos 28 famílias são colocadas no filo Archaeprotista. Os exemplos incluem Archaemoebae (Pelomyxa e Mastigamoeba), Metamonadas (Retortamonas), Diplomonads (Giardia), Oxymonadas (Pyrsonympha) e as duas ordens de Parabasalia; Trichomonadida (com Devescovina, Mixotricha, Monocercomonas, Trichomonas, Calonymphida (Coronympha, Snyderella) e Hypermastigida (Lophomonas, Staurojoenina, e Trichonympha) (Margulis et al, 2000).
Durante a evolução destes protistas, os núcleos foram retirados a partirdos cariomastigontes e acariomastigontes foram gerados (Kirby, 1949).
Núcleos desapegado temporariamente (ou não) a associados ondulipódios ou foram libertados, se proliferaram ocupando posições centrais nas células. Os ondulipódios, também geraram grandes células, onde os nadadores mais rápidos sobreviviam a esta etapa evolutiva (Margulis et al, 2000).
Outra evidência dessa relação com os eucariotos é que nos cariomastigontes o citoesqueleto é visível o centro dos membros basais, a mesma conformação encontrada em quase todas as linhagens Archaeprotistas (Archamoeba, Metamonads, e Parabasalia) (Brugerolle, 1991).
Em tricomonas, o cariomastigonte inclui um corpo (complexo de Golgi) em Parabasalia, e coordena a formação de hidrogenossomas (inclusões delimitadas por uma membrana de tamanho de células bacterianas que geram hidrogênio).
O cariomastigonte reproduz-se como uma estrutura unitária. Normalmente, quatro cinetossomos anexados com a estruturas laminadas de microtúbulos (o axostilo e suas extensões, os peltas) reproduzem-se e suas relações morfológicas são mantidas (Margulis et al, 2000).
Cinetossomos reproduzem-se primeiro, o núcleo se divide, e os dois grupos de cinetossomos separaram-se nos pólos de um eixo de microtúbulos finos chamado a paradesmose. Os cinetossomos e estruturas associadas encontram-se e repartem. Uma parte origina novos cariomastigontes integros, a outra produz componentes que faltam, tais como o complexo de Golgi e axostilo.
Genes nucleares de α-proteobacterial foram vistos como tendo se originado de mitocôndrias perdidas ou degeneradas em pelo menos duas espécies, Archaeprotistas (Giardia lamblia e Trichomonas vaginalis)(Roger et al, 1998 & Roger et al, 1996 & Germot et al, 1996) e em um microsporídio (Sogin, 1997).
Hidrogenossomas, ou pelo menos alguns tipos deles, compartilham uma origem comum com mitocôndrias. Na “hipótese do hidrogênio” (Martin & Muller, 1998), hidrogenossomas são reivindicados serem a fonte de genes eubacterianas em a-mitocondriados. Genes nucleares de Eubacterianas que não eram originários de Spirochete provavelmente foram adquiridos de protistas a-mitocondriados e de outros que não os da linhagem de simbiontes mitocondrial proteobacterial.
Bactérias Gram-negativas, que podiam estar relacionadas a ancestrais de hidrogenossomas foram epibiontes, endobiontes ou até mesmo endonuclear em Caduceia versatilis (d’Ambrosio et al, 1999).
Cariomastigontes com núcleos livres formaram-se independente em muitas linhagens tanto antes quanto após a aquisição de mitocôndrias.
O ancestrais Calonymphid de Snyderella sp tornaram seu núcleos livres antes da simbiose mitocondrial (Dolan et al, 2000), e ancestrais de Chlamydomonas e de outras clorofíceas como Acetabularia lançaram os núcleos após a linhagem se tornar totalmente aeróbia (Hall & & Luck, 1995). Em formas tróficas dos protistas que carecem de estágios mastigonte, o cariomastigonte é geralmente ausente. A única exceção é Histomonas, uma célula tricomonada amebóide que carece de um axonema mas carrega o suficiente da estrutura cariomastigonte remanescente de tal forma que é possivel classifica-la como Parabasalia ao invés de uma ameba rizópoda (Dyer, 1990).
Este sistema organelar aparece nos zoósporos, formas tróficas móveis do esperma de muitos organismos e sugere certa facilidade de desenvolvimento dos cariomastigonte (Margulis et al, 2000).
O cariomastigonte em algumas células é facilmente perdido, suprimido ou mesmo recuperado. Em muitos taxa multinucleados ou protistas multicelulares (foraminíferos, algas verdes) e até mesmo em plantas, o cariomastigonte persiste apenas em zoósporos ou gâmetas.
Em levedura, vermes nematóides, insetos e células de mamíferos os centros de organização de microtúbulos não-cariomastigontes são necessários para posicionar núcleos em localizações específicas no citoplasma (Raff, 1999). A ligação entre o centro da organização dos microtúbulos e os núcleos ainda não é inteiramente compreendida. Ao que parece, tal ligação é um legado evolutivo, um remanescente do conector archaebacterial-eubacterial original.
As organelas modernas (centríolo, cinetossomos, núcleos, Golgi, e axostilos) derivam do que primeiro assegurou a continuidade genética dos componentes de um quimera muito antigo e cariomastigonte é uma estrutura que teria sido muito mais visível no Proterozóico do é que para nós.
Victor Rossetti
Palavras chave: NetNature, Rossetti, Archezoa, Parabasalia, Archamoeba, Microsporidia, Metamonadas, Eucariogênese, Chaperonas, Simbiose, Nucleo Celular, Cariomastigonta, Lynn Margulis.
.
Referências
